
สู่รุ่นหนึ่งคือดีเอ็นเอ (DNA) ซึ่งโดยทั่วไปอยู่ในสภาพสายคู่และวางตัวเป็นเกลียวเวียนขวา

รูปที่ 1.1 โครงสร้างของดีเอ็นเอ
: DNA มีขนาดเส้นผ่าศูนย์กลางประมาณ 2 นาโนเมตรและยาวในระดับเมตร
เราจะพับ DNA อย่างไรให้ได้โครโมโซมซึ่งมีขนาดเส้นผ่าศูนย์กลางประมาณ
1000 นาโนเมตร และยาวในระดับไมโครเมตร
ในแต่ละเซลล์ของสิ่งมีชีวิตดีเอ็นเอมีความยาวเป็นเมตร ดังนั้นหากวางอยู่ในนิวเคลียสใน
ลักษณะที่พันกันอยู่อย่างไม่เป็นระเบียบ คงเป็นการยากที่จะเก็บดีเอ็นเอที่ยาวขนาดนั้นไว้ใน
ขอบเขตของนิวเคลียสที่เล็กมาก และถ้าเกิดการแบ่งเซลล์ในขณะที่ดีเอ็นเอยังคงเป็นสายยาว
และพันกันอยู่อย่างยุ่งเหยิงแล้ว ในช่วงที่แยกสารพันธุกรรมให้เซลล์ใหม่ทั้งสองเซลล์จะต้อง
เกิดการขาดของสายดีเอ็นเออย่างแน่นอน ด้วยเหตุนี้เซลล์จึงต้องมีระบบจัดเก็บสารพันธุกรรม
ให้เป็นระเบียบ โดยเริ่มจาก
ส่วนใหญ่เป็นโปรตีนฮีสโตน (histone protein) กลายเป็นโครงสร้างที่เรียกว่า โครมาทิน
(chromatin) ซึ่งมีขนาดเส้นผ่าศูนย์กลางประมาณ 10 นาโนเมตร โครงสร้างนี้เกิดจากการที่
สายดีเอ็นเอพันรอบโปรตีนฮีสโตน 8 อัน เรียกโครงสร้างที่เกิดจากการพันกันนี้ว่า
นิวคลีโอโซม (nucleosome) ในสายโครมาทิน 1 สาย จะประกอบด้วยนิวคลีโอโซม
จำนวนมาก (polynucleosomes) Roger Kornberg (1974) ศึกษาโครงสร้างของโครมาทิน
โดยใช้กล้องจุลทรรศน์อิเล็กตรอนพบว่ามีเม็ดกลมๆ จำนวนมาก และแต่ละเม็ดเชื่อมต่อกัน
ด้วยเส้นใยขนาดเล็ก นอกจากนี้เขายังได้ทดลองใช้เอ็นไซม์สำหรับตัดดีเอ็นเอมาตัดโครมาทิน
พบว่าได้ชิ้นดีเอ็นเอที่มีขนาดประมาณ 200 คู่เบส จำนวนมาก ในขณะที่เมื่อใช้เอ็นไซม์
ชนิดเดียวกันตัดสายดีเอ็นเอจะได้ชิ้นดีเอ็นเอที่มีขนาดแตกต่างกัน เขาสรุปว่าการที่ตัด
โครมาทินแล้วได้ชิ้นขนาดประมาณ 200 คู่เบสเนื่องจากเอ็นไซม์ตัดดีเอ็นเอได้เฉพาะบริเวณ
ที่อยู่ระหว่างนิวคลีโอโซม ส่วนบริเวณที่เป็นนิวคลีโอโซมเอ็นไซม์ไม่สามารถตัดสายดีเอ็นเอได้
เนื่องจากการปกป้องหรือกีดขวางโดยโปรตีนฮีสโตน
โครงสร้างที่มีเส้นผ่าศูนย์กลางใหญ่ขึ้นเรียกว่า โซลีนอยด์ (solenoid) ระยะของการขดตัว
ดังกล่าวจะยังเห็นรูปร่างไม่ชัดเจน จนกระทั่งขดตัวแน่นที่สุดจะเห็นเป็นรูปร่างคล้ายกากบาท (X) ระยะนี้จึงเรียกว่า โครโมโซม (โครโมโซมซึ่งมีรูปร่างคล้ายกากบาทนี้ จะสังเกตเห็นได้
อย่างชัดเจนในระยะเมทาเฟสของการแบ่งเซลล์ โดยโครโมโซมที่มีรูปร่างดังกล่าวประกอบ
ไปด้วยโครมาทิน 2 เส้น ติดกันอยู่ที่เซนโทรเมียร์)

รูปที่ 1.2 การจับของ DNA กับโปรตีนฮีสโตน
 |
|
: คุณคิดว่าฮอมอโลกัสโครโมโซม หรือบางครั้งเรียกว่าโครโมโซมคู่เหมือนนั้น เหมือนกันจริงหรือไม่อย่างไร |
ของโครโมโซมแต่ละคู่โดยดูจากขนาด ตำแหน่งของเซนโทรเมียร์ (centromere) และรูปแบบ
การย้อมติดสีที่แตกต่างกัน โครโมโซมซึ่งมีลักษณะเหมือนกันทั้งในด้านขนาด ตำแหน่งของ
เซนโทรเมียร์ และรูปแบบการย้อมติดสีเรียกว่า ฮอมอโลกัสโครโมโซม (homologous chromosome) อย่างไรก็ตามก็มีข้อยกเว้นสำหรับฮอมอโลกัสโครโมโซมบางคู่ ที่ไม่ได้มี
ลักษณะเหมือนกันทั้งในด้านขนาด ตำแหน่งของเซนโทรเมียร์ ตลอดจนรูปแบบการย้อมติดสี
เช่น คู่ของโครโมโซม X และ โครโมโซม Y ซึ่งเป็นโครโมโซมเพศ (sex chromosomes)
ที่พบในเซลล์มนุษย์
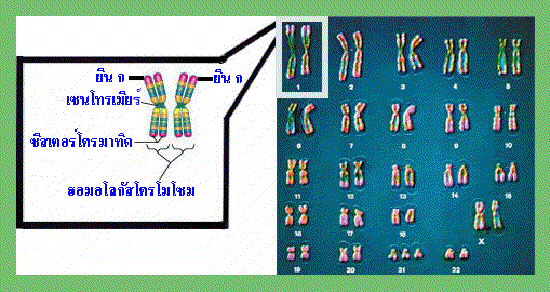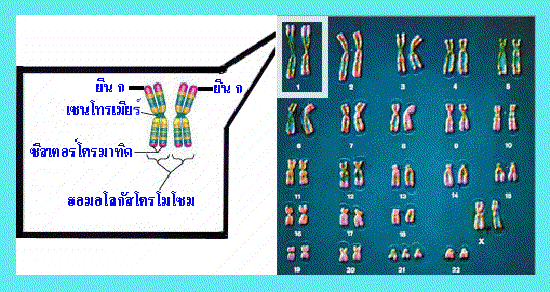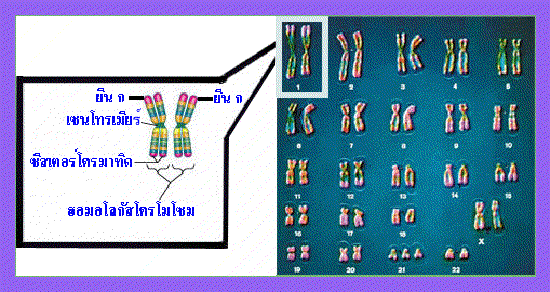
รูปที่ 1.3 ภาพแสดง homologous chromosome
การถ่ายทอดมาจากแม่ โครโมโซมทั้ง 2 แท่งนี้แม้จะมีลักษณะเหมือนกันเมื่อมองผ่าน
กล้องจุลทรรศน์ แต่กลับมีความแตกต่างของยีนที่อยู่บนตำแหน่งเดียวกัน ทำให้นอน-ซีสเตอร์
โครมาทิด (non-sister chromatids) มีความแตกต่างกันไปด้วย เช่น จากภาพด้านบนกำหนด
ให้ยีนที่อยู่บนตำแหน่ง ก ของโครโมโซมทั้ง 2 แท่ง ควบคุมลักษณะเส้นผม ยีนที่ตำแหน่ง ก
ของโครโมโซมแท่งซ้ายมือทำให้มีลักษณะผมหยัก ส่วนยีนที่ตำแหน่ง ก ของโครโมโซมแท่ง
ขวามือทำให้มีลักษณะผมตรง เป็นต้น ดังนั้นเมื่อเกิดการแลกเปลี่ยนชิ้นส่วนของสารพันธุกรรม
ระหว่างนอน-ซีสเตอร์โครมา-ทิดในการแบ่งเซลล์แบบไมโอซีส จึงทำเกิดความแปรผันของ
ลักษณะพันธุกรรมของเซลล์ใหม่ที่ได้จากการแบ่ง
|
: โครโมโซมและโครมาทินต่างกันอย่างไร : คุณคิดว่า ถ้าเกิดการแลกเปลี่ยนชิ้นส่วนของสารพันธุกรรมระหว่างซีสเตอร์ |
 |
 |