
จากที่กล่าวไปแล้วในตอนต้นว่า กระบวนการเริ่มต้นที่จะนำข้อมูลทางพันธุกรรมที่เก็บ
อยู่ในนิวเคลียสออกมาใช้ให้เกิดประโยชน์ หรือการแสดงออกถึงหน้าที่ที่เป็นรูปธรรมของยีน
บนสายดีเอ็นเอนั้น เกิดโดยผ่านกระบวนที่เรียกว่า การคัดลอกดีเอ็นเอ (DNA
transcription)
ซึ่งเป็นการสังเคราะห์อาร์เอ็นเอโดยอาศัยดีเอ็นเอเป็นแม่แบบ ด้วยเหตุนี้ข้อมูลบนสาย
อาร์เอ็นเอที่ถูกสังเคราะห์ขึ้นจึงมีความหมายเดียวกับข้อมูลบนสายดีเอ็นเอที่คอมพลีเมนทารี
(complementary) กับสายที่ถูกคัดลอก แต่อาจมีรูปลักษณ์ที่ต่างไปจากเดิม ตัวอย่างเช่น

รูปที่ 3.2 เปรียบเทียบความหมายของtranscription และtranslation
การคัดลอกดีเอ็นเอหรือการสังเคราะห์อาร์เอ็นเอทั้งในโปรแคริโอตและยูแคริโอต
เป็น
กระบวนการที่เกิดขึ้นแบบเลือก หมายความว่า จะมีเพียงบางส่วนของลำดับนิวคลีโอไทด์
บนสายดีเอ็นเอเท่านั้นที่จะถูกคัดลอกเพื่อสังเคราะห์เป็นอาร์เอ็นเอ และสำหรับในยูแคริโอต
จะมีเพียงบางส่วนของลำดับนิวคลีโอไทด์บนสายอาร์เอ็นเอ (ที่สังเคราะห์ขึ้นในตอนแรก)
เท่านั้นที่จะเข้าสู่กระบวนการแปลรหัสเพื่อสังเคราะห์เป็นโปรตีน ข้อมูลข้างต้นนี้ชี้ให้เห็นว่า
สายอาร์เอ็นเอที่ถูกสังเคราะห์ขึ้นในตอนแรกจะถูกปรับแต่ง (modified) ในบางส่วนก่อนที่จะนำ
ไปเป็นแม่แบบในการสังเคราะห์โปรตีน ซึ่งจะได้กล่าวถึงรายละเอียดในลำดับต่อไป
กระบวนการที่เกิดขึ้นแบบเลือก หมายความว่า จะมีเพียงบางส่วนของลำดับนิวคลีโอไทด์
บนสายดีเอ็นเอเท่านั้นที่จะถูกคัดลอกเพื่อสังเคราะห์เป็นอาร์เอ็นเอ และสำหรับในยูแคริโอต
จะมีเพียงบางส่วนของลำดับนิวคลีโอไทด์บนสายอาร์เอ็นเอ (ที่สังเคราะห์ขึ้นในตอนแรก)
เท่านั้นที่จะเข้าสู่กระบวนการแปลรหัสเพื่อสังเคราะห์เป็นโปรตีน ข้อมูลข้างต้นนี้ชี้ให้เห็นว่า
สายอาร์เอ็นเอที่ถูกสังเคราะห์ขึ้นในตอนแรกจะถูกปรับแต่ง (modified) ในบางส่วนก่อนที่จะนำ
ไปเป็นแม่แบบในการสังเคราะห์โปรตีน ซึ่งจะได้กล่าวถึงรายละเอียดในลำดับต่อไป
(mRNA) ribosomal RNA (rRNA) และ transfer RNA (tRNA) อาร์เอ็นเอทั้ง 3 ชนิดนี้มี
หน้าที่แตกต่างกันออกไปกล่าวคือ mRNA ทำหน้าที่เป็นแม่แบบสำหรับการสังเคราะห์โปรตีน
rRNA ทำหน้าที่เป็นส่วนประกอบของไรโบโซม และ tRNA มีหน้าที่ในการนำกรดอะมิโนอิสระ
เข้ามาต่อกันในขั้นตอนของการสังเคราะห์โปรตีน แม้ว่าจะมีหน้าที่แตกต่างกันอย่างชัดเจน
อาร์เอ็นเอทั้ง 3 ชนิดนี้ก็ยังต้องทำงานร่วมกันอย่างเป็นขบวนการ ทั้งนี้เพื่อให้การสังเคราะห์
โปรตีนเกิดขึ้นได้อย่างสมบูรณ์
เช่นเดียวกับการลอกแบบเพื่อสังเคราะห์ดีเอ็นเอเส้นใหม่ การคัดลอกดีเอ็นเอเพื่อสร้าง
เป็นอาร์เอ็นเอนั้นจำเป็นจะต้องมีการง้างเกลียวของสายดีเอ็นเอเกลียวคู่ให้แยกออกจากกัน
และใช้หลักการของการเกิดเบสคู่สม (complementary) ดังนั้นลำดับเบสบนสายอาร์เอ็นเอ
จะมีลักษณะเป็นคู่สมกันกับลำดับเบสบนดีเอ็นเอสายแม่แบบ แต่การสังเคราะห์อาร์เอ็นเอ
จะใช้ไรโบนิวคลีโอไทด์เป็นสับสเตรท ในขณะที่การสังเคราะห์ดีเอ็นเอสับสเตรทที่ใช้คือ
ดีออกซีไรโบนิวคลีโอไทด์ นอกจากนี้การเริ่มต้นของการสังเคราะห์อาร์เอ็นเอยังไม่มี
ความจำเป็นที่จะต้องใช้ไพรเมอร์ (primer) ทั้งนี้เนื่องจากเอ็นไซม์ที่สำคัญที่ใช้ในกระบวนการ
สังเคราะห์อาร์เอ็นเอที่ชื่อว่า อาร์เอ็นเอโพลีเมอเรส (RNA polymerase) มีความสามารถ
ที่จะเริ่มทำงานได้เอง ซึ่งคุณสมบัตินี้ไม่พบในเอ็นไซม์ดีเอ็นเอโพลีเมอเรส
(DNA polymerase) ซึ่งเป็นเอ็นไซม์ที่ใช้ในการสังเคราะห์ดีเอ็นเอ
 |
: จากข้อมูลข้างต้นคุณคิดว่า การสังเคราะห์ดีเอ็นเอกับการสังเคราะห์อาร์เอ็นเอ มีความคล้ายคลึงกันและแตกต่างกันในประเด็นใดบ้าง |
สิ่งจำเป็นเบื้องต้นที่จะต้องมีได้แก่ สายดีเอ็นเอ เอ็นไซม์อาร์เอ็นเอโพลีเมอเรส โปรตีนที่ช่วย
ในการสังเคราะห์อาร์เอ็นเอ และไรโบนิวคลีโอไทด์ซึ่งทำหน้าที่เป็นสับสเตรท ในส่วนของ
สายดีเอ็นเอนั้นเบื้องต้นอยู่ในสภาพสายคู่ที่พันกันเป็นเกลียว เมื่อถูกง้างเกลียวออกในช่วงที่มี
การสังเคราะห์อาร์เอ็นเอจะใช้สายใดสายหนึ่งเป็นแม่แบบซึ่งสายที่ถูกใช้เรียกว่า เทมเพลท
(template) ส่วนอีกสายหนึ่งที่ไม่ได้ใช้เป็นแม่แบบเรียกว่า coding ซึ่งอาร์เอ็นเอที่
สังเคราะห์ได้จะมีลำดับเบสคล้ายกับลำดับเบสบนสาย coding เพียงแต่เปลี่ยนจากเบส T
เป็นเบส U เท่านั้น
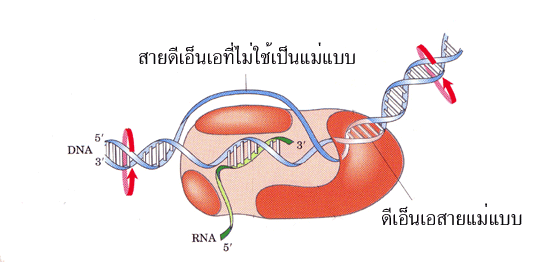
รูปที่ 3.3 การสร้าง RNA จาก DNA สายแม่แบบ
|
: คุณคิดว่าสายดีเอ็นเอที่พับซ้อนกันแน่นในโครโมโซมจะคลายเกลียว เพื่อให้เกิดการสังเคราะห์อาร์เอ็นเอได้อย่างไร : คุณมีความคิดเห็นอย่างไรกับคำกล่าวที่ว่า “มีดีเอ็นเอเพียงสายเดียวเท่านั้น (จากทั้งหมด 2 สาย) ที่สามารถทำหน้าที่เป็นแม่แบบเพื่อการสังเคราะห์ อาร์เอ็นเอได้” |
การคัดลอกดีเอ็นเอเพื่อสร้างเป็นอาร์เอ็นเอได้แก่ โปรโมเตอร์ (promoter) ซึ่งเป็นตำแหน่ง
ที่จะให้เอ็นไซม์อาร์เอ็นเอโพลีเมอเรสมาจับเพื่อที่จะเริ่มต้นสังเคราะห์อาร์เอ็นเอต่อไป
ในแบคทีเรียชนิด Escherichia coli ซึ่งจัดเป็นโปรแคริโอตชนิดหนึ่ง จะมีโปรโมเตอร์ประกอบ
ไปด้วยลำดับนิวคลีโอไทด์ 2 ชุดๆ ละ 6 นิวคลีโอไทด์ ดังภาพ

รูปที่ 3.3 โปรโมเตอร์ของยีนใน E.coli
start site) มีสัญลักษณ์เป็น +1 นิวคลีโอไทด์ชุดแรกของโปรโมเตอร์จะอยู่ห่างจาก
ตำแหน่งนี้ออกไปทางด้าน 3' (ด้านที่อยู่ก่อนที่จะถึงตำแหน่งที่จะเริ่มการสังเคราะห์อาร์เอ็นเอ)
ของสายแม่แบบ 10 คู่เบส ให้สัญลักษณ์ของตำแหน่งนี้เป็น -10 (- เป็นสัญลักษณ์ว่าอยู่ด้านที่
อยู่ก่อนที่จะถึงตำแหน่งที่จะเริ่มการสังเคราะห์อาร์เอ็นเอ ส่วน 10 เป็นสัญลักษณ์ของระยะห่าง
จากตำแหน่งที่จะเริ่มการสังเคราะห์อาร์เอ็นเอ) นิวคลีโอไทด์ชุดที่สองของโปรโมเตอร์จะอยู่ที่
ตำแหน่ง -35 ลำดับเบสในนิวคลีโอไทด์ทั้งสองชุดของโปรโมเตอร์ของยีนที่ต่างกันจะคล้ายกัน
มาก (อาจมีการสลับตำแหน่งกันบ้างเท่านั้น) แต่โดยส่วนใหญ่แล้วจะมีลำดับดังแสดงไว้ใน
รูปที่ 3.3
การทำงานของเอ็นไซม์อาร์เอ็นเอโพลีเมอเรสทั้ง 3 ชนิด โดยโปรโมเตอร์ที่สำคัญของยีนที่ถูก
คัดลอกโดยเอ็นไซม์ RNA polymerase II มีชื่อเรียกว่า TATA box (เพราะมีลำดับเบสเป็น
TATAA) มีตำแหน่งอยู่ที่ -25 ถึง -30 ของตำแหน่งที่จะเริ่มการสังเคราะห์ mRNA โดย TATA
box หากเปรียบไปแล้วก็คงมีศักดิ์เทียบเท่ากับตำแหน่งโปรโมเตอร์ที่ -10 ของ E. coli นั่นเอง
สำหรับโปรโมเตอร์ของยีนที่ถูกคัดลอกโดยเอ็นไซม์ RNA polymerase I จะครอบคลุมพื้นที่
ตั้งแต่ตำแหน่งที่จะเริ่มการสังเคราะห์อาร์เอ็นเอไปจนถึงตำแหน่ง -150 ส่วนโปรโมเตอร์ของยีน
ที่ถูกคัดลอกโดยเอ็นไซม์ RNA polymerase III ส่วนใหญ่จะมีลำดับนิวคลีโอไทด์อยู่ใน
ส่วนต้นของยีนที่จะถูกคัดลอกนั่นเอง
|
: ถ้าตำแหน่งโปรโมเตอร์เกิดความเสียหาย หรือเอ็นไซม์อาร์เอ็นเอโพลีเมอเรส ไม่สามารถจดจำตำแหน่งของโปรโมเตอร์ได้จะส่งผลอย่างไรต่อการ การคัดลอกดีเอ็นเอเพื่อสร้างอาร์เอ็นเอหรือไม่ |
สังเคราะห์เป็นสายอาร์เอ็นเอโดยการนำนิวคลีโอไทด์ตัวใหม่เข้าไปต่อที่ปลายด้าน 3'
ของสายอาร์เอ็นเอ (ทำให้โมเลกุลของอาร์เอ็นเอยืดยาวออกไปในทิศ 5' ไป 3') นั้นมี
ความแตกต่างกันทั้งในด้านองค์ประกอบของโครงสร้าง และคุณสมบัติระหว่างของ
โปรแคริโอต และยูแคริโอต จึงแยกอธิบายดังนี้ เอ็นไซม์อาร์เอ็นเอโพลีเมอเรสใน
โปรแคริโอตมีอยู่เพียงชนิดเดียว ประกอบขึ้นด้วยโพลีเพปไทด์หลายสาย และมีหน่วยย่อย
(subunit) ที่ต่างกัน 5 หน่วย ได้แก่
ด้วยสีเขียวและสีเขียวอ่อน),
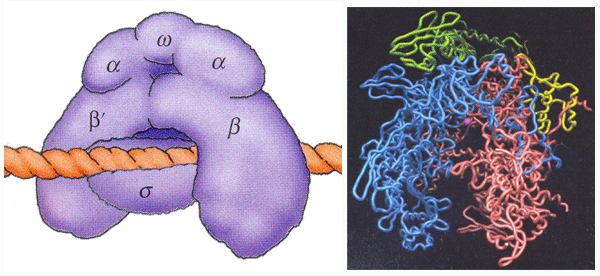
รูปที่ 3.4 เอ็นไซม์อาร์เอ็นเอโพลีเมอเรสในโปรแคริโอต
หน่วยย่อยอื่นได้ภายหลังจากที่มีสังเคราะห์อาร์เอ็นเอไปได้ประมาณ 8-10 นิวคลีโอไทด์
เมื่อหน่วยย่อยนี้หลุดออกไป เอ็นไซม์อาร์เอ็นเอโพลีเมอเรสยังคงทำหน้าที่ต่อไปได้อย่างปกติ
จึงเป็นการชี้ให้เห็นว่า หน่วยย่อย
เอ็นไซม์ชนิดนี้ อย่างไรก็ตามพบว่า เอ็นไซม์อาร์เอ็นเอโพลีเมอเรสจะไม่สามารถจับกับ
สายดีเอ็นเอตรงตำแหน่งที่จำเพาะที่เรียกว่า โปรโมเตอร์ได้หากปราศจากความช่วยเหลือ
ของหน่วยย่อย
ของลำดับเบสที่ถูกต้อง เพื่อนำพาเอ็นไซม์อาร์เอ็นเอโพลีเมอเรสมาจับกับสายดีเอ็นเอ
เพื่อเริ่มต้นการสังเคราะห์อาร์เอ็นเอต่อไป
ชนิดทำหน้าที่ในการสังเคราะห์อาร์เอ็นเอต่างชนิดกัน กล่าวคือ RNA polymerase I ทำหน้าที่
ในการสังเคราะห์ rRNA (5.8S, 18S, 28S) RNA polymerase II ทำหน้าที่ในการสังเคราะห์
mRNA และ RNA polymerase III ทำหน้าที่ในการสังเคราะห์ tRNA และ rRNA (5S)
(ค่า S ของ rRNA แปรผันตรงกับขนาดของ rRNA หมายความว่าค่า S ยิ่งมากrRNA ยิ่งมี
ขนาดใหญ่นั่นเอง) เอ็นไซม์ทั้ง 3 ชนิดนี้ประกอบขึ้นด้วยหน่วยย่อย 12-17 หน่วย เอ็นไซม์
อาร์เอ็นเอโพลีเมอเรสในยูแคริโอต จะไม่สามารถไปจับกับสายดีเอ็นเอได้โดยตรงเหมือนเช่น
ในโปรแคริโอต แต่จำเป็นจะต้องอาศัยความช่วยเหลือของโปรตีนที่ทำหน้าที่เป็น
ทรานสคริปชันแฟคเตอร์ (transcription factor) อีกหลายตัว ซึ่งจะกล่าวถึงรายละเอียด
ในลำดับต่อไป
อาร์เอ็นเอโพลีเมอเรสแต่ละชนิดอยู่ประมาณ 20,000 ถึง 40,000 โมเลกุล โดยปริมาณของ
เอ็นไซม์นี้จะถูกควบคุมให้มีความสัมพันธ์กับอัตราการเจริญเติบโตของเซลล์
|
: คุณคิดว่า ในระยะใดของการแบ่งเซลล์ที่เอ็นไซม์อาร์เอ็นเอโพลีเมอเรส มีกิจกรรมการทำงานสูงสุด และต่ำสุด เพราะเหตุใด |

