
ลักษณะเหมือนกันคือสามารถแบ่งออกได้เป็น 3 ขั้นตอนใหญ่ๆ ได้แก่ ขั้นเริ่มต้น (initiation)
ขั้นต่อสาย (elongation) และขั้นหยุด (termination)
|
ขั้นเริ่มต้น |
เป็นขั้นที่เอ็นไซม์อาร์เอ็นเอโพลีเมอเรสมาจับกับสายดีเอ็นตรงตำแหน่ง
โปรโมเตอร์ และง้างเกลียวคู่ของสายดีเอ็นเอให้แยกออกจากกัน โดยในโปรแคริโอต
เอ็นไซม์อาร์เอ็นเอ
โพลีเมอเรสจะเข้าจับกับสายดีเอ็นเอตรงบริเวณก่อนที่จะถึงตำแหน่งของโปรโมเตอร์
(upstream ของโปรโมเตอร์) และจะเคลื่อนที่มาเรื่อยๆ ตามสายดีเอ็นเอจนกระทั่งมาถึง
ตำแหน่งโปรโมเตอร์ที่ถูกจดจำโดยหน่วยย่อย ![]() เอ็นไซม์ก็จะหยุด (ขนาดของเอ็นไซม์มีพื้นที่
เอ็นไซม์ก็จะหยุด (ขนาดของเอ็นไซม์มีพื้นที่
ครอบคลุมทั้งตำแหน่งของโปรโมเตอร์ และตำแหน่งเริ่มต้นของการสังเคราะห์อาร์เอ็นเอ)
และเอ็นไซม์จะง้างเกลียวของดีเอ็นเอให้แยกออกจากกันประมาณ 14-17 คู่เบส
(จากตำแหน่ง
+2 ถึง -12) การง้างเกลียวนี้เป็นผลให้ดีเอ็นเออยู่ในสภาพพร้อมที่จะเป็นแม่แบบสำหรับ
การสังเคราะห์อาร์เอ็นเอในขั้นตอนต่อไป

รูปที่ 3.5 ขั้นเริ่มต้น (initiation) ของการสังเคราะห์ RNA ในโปรแคริโอต
โพลีเมอเรสแต่โปรตีนที่ทำหน้าที่เป็นทรานสคริปชันแฟคเตอร์อีกหลายชนิด ดังนั้นในขั้นเริ่มต้น
จึงเป็นการเข้าจับของทรานสคริปชันแฟคเตอร์ตรงตำแหน่งโปรโมเตอร์ก่อน แล้วจึงเหนี่ยวนำ
ให้มีการเข้าจับของเอ็นไซม์อาร์เอนเอโพลีเมอเรสตรงตำแหน่งโปรโมเตอร์อีกทีหนึ่ง จากนั้นก็
ยังมีการเข้ามาจับสายดีเอ็นเอของทรานสคริปชันแฟคเตอร์อีกหลายชนิด (โดยตัวที่เข้ามาตอน
ท้ายนี้จะเป็นตัวช่วยให้เอ็นไซม์อาร์เอ็นเอโพลีเมอเรสเคลื่อนที่ออกจากจุดเริ่มต้น เพื่อทำการ
สังเคราะห์อาร์เอ็นเอได้) เมื่อผ่านขั้นตอนนี้แล้วเอ็นไซม์จึงจะง้างเกลียวของดีเอ็นเอเพื่อให้เกิด
การทำงานในขั้นตอนต่อไป

รูปที่ 3.6 ขั้นเริ่มต้น (initiation) ของการสังเคราะห์ RNA ในยูแคริโอต
 |
: การง้างเกลียวคู่ของสายดีเอ็นจะทำให้เกิดความเครียดหรือแรงกดตรงบริเวณ เกลียวคู่ก่อนและหลังจุดที่เกิดการง้างเกลียว คุณคิดว่าเซลล์มีวิธีแก้ปัญหา เรื่องความเครียดหรือแรงกดที่เกิดขึ้นนี้อย่างไร |
| ขั้นต่อสาย |
นิวคลีโอไทด์เข้ามาต่อที่ปลายด้าน 3' ของสายอาร์เอ็นเอ (ทำให้โมเลกุลของอาร์เอ็นเอ
ยืดยาวออกไปในทิศ 5' ไป 3') ในระหว่างที่เอ็นไซม์อาร์เอ็นเอโพลีเมอเรสเคลื่อนที่ไป
เพื่อการสังเคราะห์อาร์เอ็นเอ ก็จะเกิดการง้างเกลียวของดีเอ็นเอทางด้านหน้าของเอ็นไซม์
อย่างต่อเนื่อง ส่วนดีเอ็นเอทางด้านหลังของเอ็นไซม์ซึ่งได้ผ่านการคัดลอกไปแล้วจะกลับพัน
เป็นเกลียวเหมือนเดิม อย่างไรก็ตามจะยังคงมีดีเอ็นเอที่อยู่ในสภาพคลายเกลียวอยู่ประมาณ
15- 17 คู่เบสเสมอ เนื่องจากยีนต่างๆ ที่ใช้เป็นแม่แบบในการคัดลอกมีตำแหน่งกระจายตัว
อยู่บนสายดีเอ็นเอ และมีโปรโมเตอร์จำเพาะของตัวเอง ดังนั้นการสังเคราะห์อาร์เอ็นเอ
จึงสามารถเกิดขึ้นพร้อมกันได้ในหลายๆ ยีนบนดีเอ็นเอโมเลกุลเดียวกัน ในขั้นต่อสายของ
ยูแคริโอต พบว่ามีโปรตีนอีกหลายชนิดที่ช่วยเพิ่มประสิทธิภาพการทำงานของเอ็นไซม์อาร์
เอ็นเอโพลีเมอเรส เรียกโปรตีนเหล่านั้นว่า อิลองเกชันแฟคเตอร์ (elongation factor)
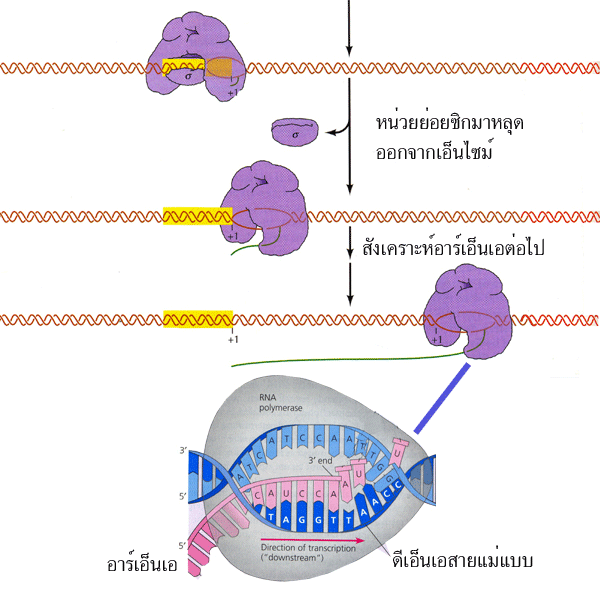
รูปที่ 3.7 ขั้นต่อสาย (elongation) ของการสังเคราะห์ RNA
|
: คุณคิดว่าพันธะที่ใช้ยึดระหว่างนิวคลีโอไทด์แต่ละตัวบนสายอาร์เอ็นเอ กับพันธะที่ใช้ยึดระหว่างนิวคลีโอไทด์บนสายดีเอ็นเอเป็นพันธะชนิดใด และเหมือนกันหรือไม่ |
| ขั้นหยุด |
โพลีเมอเรสยังสามารถนำนิวคลีโอไทด์เข้ามาต่อกับสายอาร์เอ็นเอได้ แต่การทำงานนี้
จะหยุดลงเมื่อเอ็นไซม์ได้รับสัญญาณให้หยุดหรือจบการสังเคราะห์อาร์เอ็นเอ
(termination signal)
ที่ทำงานโดยไม่ขึ้นกับการทำงานของโปรตีนที่มีชื่อว่า rho (อ่านว่า โร ใช้สัญลักษณ์
การทำงานดังนี้ ในกรณีที่การหยุดหรือจบการสังเคราะห์อาร์เอ็นเอโดยไม่ขึ้นกับการทำงานของ
โปรตีน rho จะเป็นผลมาจากลำดับเบสบนสายดีเอ็นเอแม่แบบนั่นเอง กล่าวคือ เมื่อเอ็นไซม์
เคลื่อนที่มาถึงบริเวณที่จะหยุดหรือจบการสังเคราะห์อาร์เอ็นเอ จะพบกับลำดับเบสบนสาย
ดีเอ็นเอที่มีปริมาณเบส G และ C เรียงต่อกันจำนวนมาก อาร์เอ็นเอที่เกิดจากการคัดลอก
ดีเอ็นเอบริเวณนี้จะเกิดการจับคู่กันเองระหว่างเบส G และ C ซึ่งลักษณะของการจับจะเหมือน
กับที่เกิดขึ้นบนสายดีเอ็นเอ ทำให้มีลักษณะเป็นรูปห่วง (loop) ถัดจากลำดับเบสเหล่านี้จะต่อ
ด้วยลำดับเบส A เรียงต่อกันอีกจำนวนมากอาร์เอ็นเอที่เกิดจากการคัดลอกดีเอ็นเอบริเวณนี้
(มีเบส U ต่อกันจำนวนมาก) จะมีความสามารถในการเกาะกับสายดีเอ็นเอแม่แบบได้ไม่ดีนัก
(เพราะเป็นเพียงพันธะไฮโดรเจนระหว่างเบส A-U) การจับคู่กันเองระหว่างเบส G และ C จน
ทำให้อาร์เอ็นเอมีลักษณะเป็นรูปห่วงนั้นจะส่งผลให้แรงยึดระหว่างสายอาร์เอ็นเอกับเอ็นไซม์
อาร์เอ็นเอโพลีเมอเรสมีน้อยลง ซึ่งจะกระตุ้นให้สายอาร์เอ็นเอหลุดออกจากเอ็นไซม์อาร์เอ็นเอ
โพลีเมอเรสได้ง่ายขึ้น เมื่อบวกกับพันธะที่จับกันอย่างอ่อนๆ ระหว่างเบส A-U แล้ว ผลรวมที่
เกิดตามมาคือ สายอาร์เอ็นเอ (สายเดี่ยว) จะหลุดออกจากบริเวณที่มีการคัดลอก ตามมาด้วย
การหลุดออกจากสายดีเอ็นเอของเอ็นไซม์อาร์เอ็นเอโพลีเมอเรส และท้ายที่สุดดีเอ็นเอที่อยู่ใน
สภาพคลายเกลียวก็จะกลับมาพันเกลียวกันเหมือนเดิม ในกรณีที่การหยุดหรือจบการสังเคราะห์
อาร์เอ็นเอที่ขึ้นกับการทำงานของโปรตีน rho นั้นจะพบในกรณีที่ดีเอ็นเอสายแม่แบบไม่มีลำดับ
เบส A เรียงต่อกันอีกจำนวนมาก มีเพียงเบส G และ C เรียงต่อกันจำนวนมากจนเกิดการสร้าง
รูปห่วงของสายอาร์เอ็นเอ ในกรณีนี้โปรตีน rho จะเข้าจับกับสายอาร์เอ็นเอตรงตำแหน่งจับที่
จำเพาะซึ่งอยู่ก่อนตำแหน่งที่จะเกิดการสร้างรูปห่วง จากนั้นจะเคลื่อนที่มาเรื่อยๆ จนกระทั้งมา
หยุดอยู่ที่ตำแหน่งที่เป็นรูปห่วง แล้วทำให้เกิดการหลุดออกจากกันขององค์ประกอบต่างๆ เช่น
เดียวกับในกรณีแรก แต่กลไกการทำงานที่ส่งผลให้เกิดเหตุการณ์ในลักษณะนี้ยังไม่เป็นที่ทราบ
แน่ชัด
เข้ามาต่อกับสายอาร์เอ็นเอได้อีกกว่าร้อยหน่วย แม้ว่าจะเคลื่อนที่ผ่านตำแหน่งให้หยุดหรือจบ
การสังเคราะห์อาร์เอ็นเอไปแล้ว (ได้แก่ลำดับเบส AAUAAA) ด้วยเหตุนี้สัญญาณเตือนดังกล่าว
จึงถูกคัดลอก และไปปรากฏอยู่บน mRNA แต่ถัดจากลำดับเบสนี้ไปอีกประมาณ 10-35
นิวคลีโอไทด์ mRNA นี้จะถูกตัดด้วยเอ็นไซม์ทำให้สายอาร์เอ็นเอหลุดออกจากบริเวณที่มี
การคัดลอก ตามมาด้วยการหลุดของเอ็นไซม์อาร์เอ็นเอโพลีเมอเรส ออกจากสายดีเอ็นเอ
และการคืนสู่สภาพเกลียวคู่ของสายดีเอ็นเอ
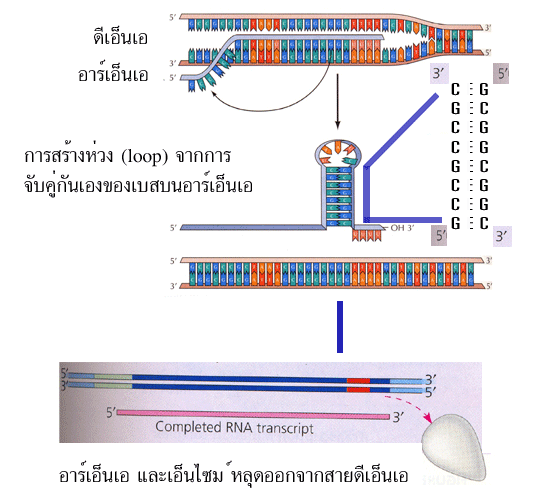
รูปที่ 3.7 ขั้นตอนหยุด (termination) ของการสังเคราะห์ RNA
ที่ยังไม่พร้อมที่จะถูกนำไปใช้งาน จึงต้องมีการตัดแต่งโมเลกุลของอาร์เอ็นเอเหล่านี้เสียก่อน
ซึ่งขั้นตอนของการตัดแต่งนั้นมากบ้างน้อยบ้างขึ้นอยู่กับชนิดของอาร์เอ็นเอ วิธีการของ
การตัดแต่งโมเลกุลของอาร์เอ็นเอชนิดต่างๆ จะกล่าวโดยละเอียดในหัวข้อถัดไป
|
: คุณคิดว่าถ้าเอ็นไซม์อาร์เอ็นเอโพลีเมอเรสหลุดออกจากสายดีเอ็นเอแม่แบบ ก่อนที่การสังเคราะห์อาร์เอ็นเอจะเสร็จสิ้น เอ็นไซม์นี้จะกลับเข้ามา ณ ตำแหน่งที่หลุดออกไปแล้วทำการสังเคราะห์อาร์เอ็นเอต่อให้เสร็จได้หรือไม่ |

