![]()
![]()
![]()
![]()
|
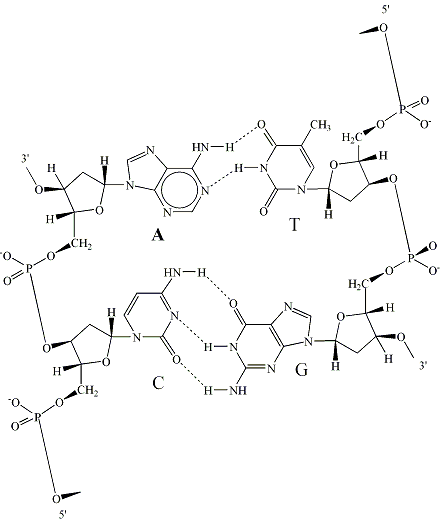
เบสอะดีนีนจะจับคู่กับไทมีน และกวานีนจะจับคู่กับไซโทซีน การจับคู่เบสแบบนี้เรียกว่า การจับคู่เบสแบบวัตสันและคริก (Watson-Crick base-pairing) ซึ่งสามารถเกิดขึ้นได้ระหว่างสายดีเอ็นเอสองสาย หรือระหว่างสายดีเอ็นเอหนึ่งสายกับสายอาร์เอ็นเอหนึ่งสาย หรือแม้แต่สายดีเอ็นเอหรืออาร์เอ็นเอสายเดียวที่พับกลับเข้าหาตัว
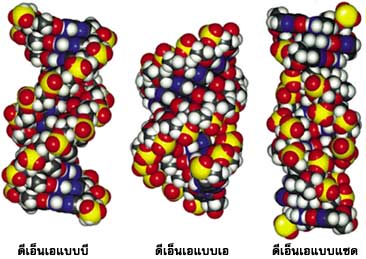
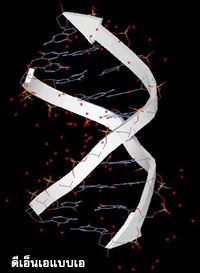
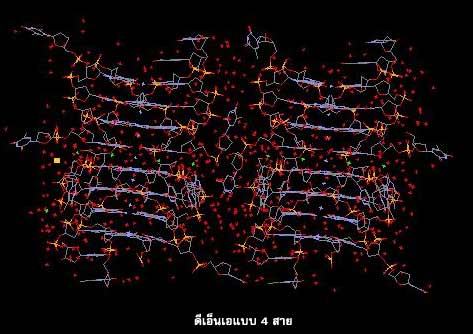
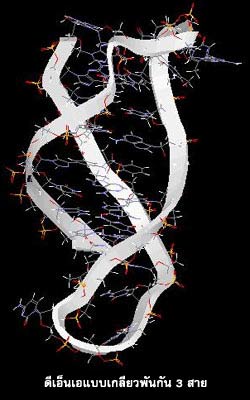
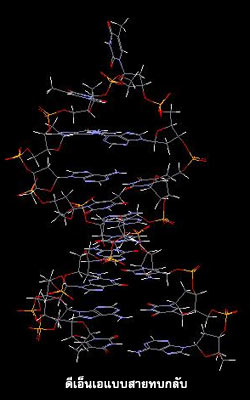
ดีเอ็นเอจะมีโครงสร้างแบบใดมักขึ้นอยู่กับชนิดของเบส ปริมาณน้ำ และอิออนโลหะที่จับอยู่กับดีเอ็นเอ โครงสร้างของดีเอ็นเอนอกจากจะเป็นชนิดสายคู่แล้ว ยังมีโครงสร้างแบบอื่นๆ เช่น ดีเอ็นเอแบบ 4 สาย ดีเอ็นเอแบบเกลียวพันกัน 3 สาย และดีเอ็นเอแบบสายทบกลับ อีกด้วย
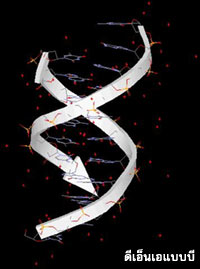
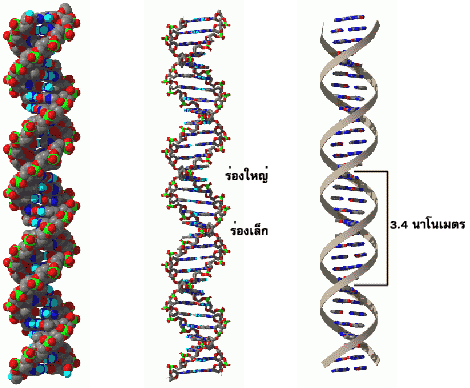
โครงสร้างของดีเอ็นเอตามแบบจำลองที่นักชีววิทยาชาวอเมริกันชื่อ เจมส์ วัตสัน (James Watson) กับนักฟิสิกส์ชาวอังกฤษชื่อ ฟรานซิส คริก (Francis Crick) ได้เสนอขึ้นในปี ค. ศ. 1953 ที่นับได้ว่าเป็นการไขปริศนาความเร้นลับของชีวิตครั้งสำคัญแห่งศตวรรษนั้น เป็นโครงสร้างของดีเอ็นเอเกลียวคู่เวียนขวาแบบบี ซึ่งมีระยะหนึ่งรอบเกลียวเท่ากับ 3.4 นาโนเมตร หรือ 34 อังสตรอม มีเบสที่เข้าคู่กัน10 คู่ต่อรอบ เกลียวนี้มีร่องใหญ่ (major groove) และร่องเล็ก (minor groove) ตลอดความยาวของเกลียว
ในการสืบพันธุ์และการแบ่งเซลล์ ไม่ว่าจะเป็นกระบวนการแบบไมโทซิส (mitosis) หรือ ไมโอซิส (meiosis) ก็ตาม ต้องมีการสร้างและจำลองแบบโครโมโซมแต่ละโครโมโซมให้เป็น 2 โครมาทิด (chromatids) ซึ่งต่างก็มีโครงสร้างและส่วนประกอบเหมือนเดิมทุกประการ ดังนั้นยีนหรือหน่วยพันธุกรรมซึ่งเป็นส่วนหนึ่งของสายดีเอ็นเอและมีการถ่ายทอดไปตามโครโมโซม ก็ต้องมีสมบัติที่สามารถสังเคราะห์ ถอดแบบ และจำลองตัวเองได้ด้วย วัตสันและคริก ได้อธิบายกลไกที่น่าเป็นไปได้เกี่ยวกับกระบวนการถอดแบบและจำลองโมเลกุลดีเอ็นเอ (DNA replication) ขึ้นมาใหม่ จาก 1 โมเลกุล เป็น 2 โมเลกุล ไว้ว่า กระบวนการดังกล่าวเริ่มต้นจากการที่สายดีเอ็นเอเกลียวคู่คลายเกลียวออก และพันธะไฮโดรเจนที่ยึดระหว่างเบสที่เข้าคู่กันสลายไปทำให้สายคู่ของดีเอ็นเอเปิดอ้าออกคล้ายกับการรูดซิปเปิด ดีเอ็นเอแต่ละสายทำหน้าที่เป็นต้นแบบ (template) สำหรับการสร้างดีเอ็นเอสายใหม่ประกบกับสายเดิม เช่น ถ้าตรงจุดหนึ่งในดีเอ็นเอสายเดิมมีนิวคลีโอไทด์ที่มีเบสเป็น T นิวคลีโอไทด์ตัวที่จะเข้ามาประกบตรงจุดนั้นจะต้องมีเบสเป็น A หรือถ้าสายเดิมมีนิวคลีโอไทด์ที่มีเบสเป็น G นิวคลีโอไทด์ตัวที่จะเข้ามาประกบตรงจุดนั้นจะต้องมีเบสเป็น C เมื่อกระบวนการถอดแบบและจำลองโมเลกุลดีเอ็นเอเสร็จสิ้นสมบูรณ์ ก็จะได้ดีเอ็นเอ 2 โมเลกุล ที่มีโครงสร้างเหมือนโมเลกุลเดิมทุกประการ โดยที่ในแต่ละโมเลกุลมีดีเอ็นเอสายเดิม 1 สาย กับสายใหม่ 1 สายเสมอ
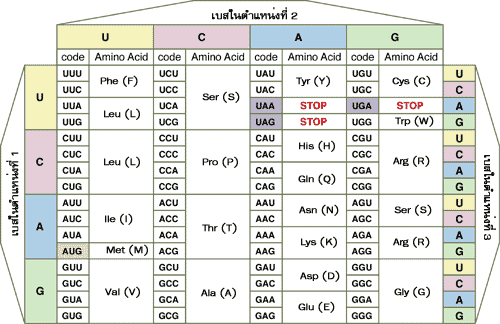
ความรู้ในทางชีววิทยาที่ว่ายีนทำหน้าที่ควบคุมลักษณะทางพันธุกรรมนั้น แสดงให้เห็นว่าการแสดงลักษณะต่างๆ ของสิ่งมีชีวิตเกิดขึ้นจากการสื่อความหมายทางพันธุกรรมจากยีนบนสายดีเอ็นเอไปสู่โปรตีน การสื่อความหมายทางพันธุกรรมจากยีนไปสู่โปรตีนอาศัยการทำงานของอาร์เอ็นเอในเซลล์ ซึ่งประกอบไปด้วยขั้นตอนการถ่ายทอดข้อมูลพันธุกรรมจากดีเอ็นเอ ที่เรียกว่า การถอดรหัส (transcription) และการเปลี่ยนรหัสจากรหัสพันธุกรรมเป็นรหัสของโปรตีน ที่เรียกว่า การแปลรหัส (translation) อาร์เอ็นเอส่วนใหญ่จะพบอยู่ในไซโทพลาซึม มีโครงสร้างเป็นสายเดี่ยวแต่อาจมีการจับคู่ของเบสภายในเส้นเดียวกันทำให้เกิดห่วงภายในโครงสร้าง เราสามารถแบ่งชนิดของอาร์เอ็นเอได้ตามรูปร่างและหน้าที่ของมันได้เป็นสามพวกใหญ่ คือ อาร์เอ็นเอนำรหัส (messenger RNA หรือ mRNA) อาร์เอ็นเอส่งถ่าย (transfer RNA หรือ tRNA) และ อาร์เอ็นเอของไรโบโซม (ribosomal RNA หรือ rRNA) อาร์เอ็นเอนำรหัสเป็นตัวรับข้อมูลพันธุกรรมจากดีเอ็นเอ โมเลกุลของมันถูกสร้างขึ้นจากดีเอ็นเอต้นแบบสายหนึ่ง จึงมีลำดับนิวคลีโอไทด์เป็นลำดับที่เข้าคู่กับสายดีเอ็นเอต้นแบบ
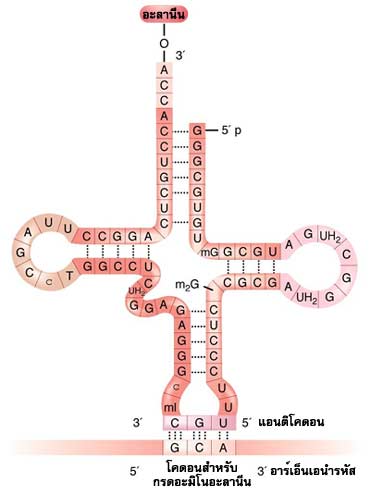
อาร์เอ็นเอส่งถ่ายมีบทบาทในขั้นตอนการแปลรหัสโดยทำหน้าที่เป็นตัวขนย้ายกรดอะมิโนไปต่อเข้าด้วยกันบนไรโบโซมจนเป็นพอลิเพปไทด์สายยาวตามลำดับรหัสกรดอะมิโนบนสายอาร์เอ็นเอนำรหัส
ในอาร์เอ็นเอส่งถ่ายแต่ละโมเลกุลมีเบสส่วนหนึ่งที่ทำหน้าที่พ่วงกรดอะมิโนได้ 1 โมเลกุล และยังมีเบสส่วนหนึ่งที่ที่สามารถจับคู่กับเบสในรหัสของอาร์เอ็นเอนำรหัส เบสส่วนนี้เรียกว่าแอนติโคดอน ประกอบไปด้วยเบส 3 ตัว ที่เข้าคู่กับโคดอนบนสายอาร์เอ็นเอนำรหัส 1 โคดอน อาร์เอ็นเอส่งถ่ายแต่ละโมเลกุลจึงมีแอนติโคดอนที่แตกต่างกัน และจะพ่วงกรดอะมิโนที่มีความสัมพันธ์กับแอนติโคดอนนั้น เช่น อาร์เอ็นเอส่งถ่าย ที่มีแอนติโคดอนประกอบไปด้วยเบส UAG ก็จะจับคู่กับโคดอน AUC ของอาร์เอ็นเอนำรหัส และจะพ่วงกรดอะมิโนไอโซลิวซีน แต่อย่างไรก็ตามการจับคู่เบสระหว่างแอนติโคดอนของอาร์เอ็นเอส่งถ่ายกับโคดอนของอาร์เอ็นเอนำรหัส อาจเป็นแบบอื่นที่ไม่ใช่การจับคู่เบสแบบวัตสันและคริกก็ได้ และในโมเลกุลของอาร์เอ็นเอส่งถ่ายในบริเวณอื่นๆ ยังอาจมีเบสชนิดอื่นที่นอกเหนือไปจากเบสพื้นฐาน 4 ตัว คือ A, U, G และC อีกด้วย ส่วนอาร์เอ็นเอของไรโบโซมช่วยในการสื่อความหมายระหว่างอาร์เอ็นเอนำรหัสและอาร์เอ็นเอส่งถ่ายที่มาประกบกันบนไรโบโซมในขณะที่มีการสร้างสายพอลิเพปไทด์ของโปรตีน
|