การคัดลอกมีกลไกคล้ายกับการลอกแบบเพื่อสร้าง DNA การคัดลอกเพื่อสร้าง RNA นั้น DNA จะต้องมีการคลายเกลียวออก และใช้หลักการของการเกิดเบสคู่สม ดังนั้นลำดับเบสบน RNA จะมีลำดับเบสที่เป็นคู่สมกันกับลำดับเบสบน DNA สายแม่แบบ แต่การสร้าง RNA จะใช้ไรโบนิวคลีโอไทด์เป็นสับสเตรท (สร้างโดยอาร์เอนเอโพลีเมอเรส) และไม่ต้องการไพรเมอร์
การคัดลอกแบบดีเอนเอ เป็นการสังเคราะห์
mRNA โดยใช้ DNA เป็นแม่แบบ ใน DNA เกลียวคู่จะใช้สายใดสายหนึ่งเป็นแม่แบบ
เรียกว่าสาย template ส่วนอีกสายที่ไม่ได้ใช้เป็นแม่แบบในการสังเคราะห์
RNA เรียกสาย coding (ภาพที่ 3.11) ขั้นตอนการสังเคราะห์ RNA แบ่งเป็น
3 ขั้นตอนคือ ขั้นเริ่มต้น (initiation) ขั้นต่อสาย (elongation) และขั้นหยุด
(termination)
![]()
ภาพที่ 3.11 แสดงการคัดลอกจาก DNA เป็น RNA
![]()
คำถาม : |
DNA ที่พับซ้อนกันแน่นในโครโมโซมจะคลายเกลียวเพื่อให้เกิดการลอกแบบและการคัดลอกได้อย่างไร |
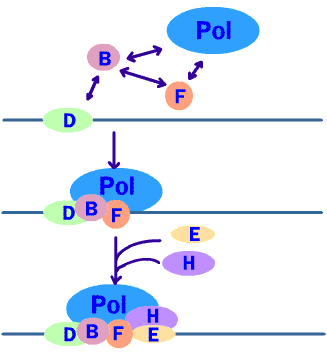
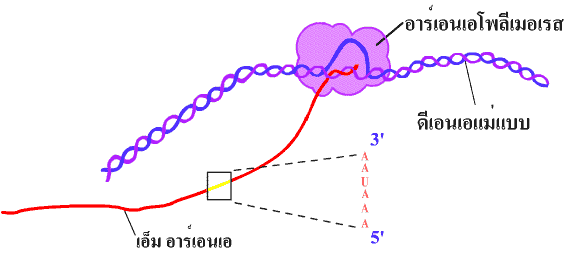
Initiation เป็นขั้นตอนเริ่มต้นที่อาร์เอนเอโพลีเมอเรส (RNA polymerase) จะมาจับกับ DNA สายแม่แบบตรงตำแหน่ง โปรโมเตอร์ (promotor) โดยอาศัยการทำงานร่วมกันของอินนิทิเอชันแฟคเตอร์ (initiation factor) หลายชนิดซึ่งจะเข้ามาเกาะบริเวณโปรโมเตอร์ และ เอนฮานเซอร์ (enhancer) ก่อน จากนั้นจึงเหนี่ยวนำให้อาร์เอนเอโพลีเมอเรสเข้ามาจับโปรโมเตอร์ได้ และทำให้เกลียวคู่ของ DNA โป่งออกบริเวณที่มีการลอกรหัส (ภาพที่ 3.12 และ 3.13)
ภาพที่
3.12 ขั้นตอนเริ่มต้นของการคัดลอกดีเอนเอโดยเอนไซม์อาร์เอนเอโพลีเมอเรส และแฟคเตอร์ต่างๆ
บริเวณโปรโมเตอร์ของ DNA อย่างเป็นลำดับขั้น
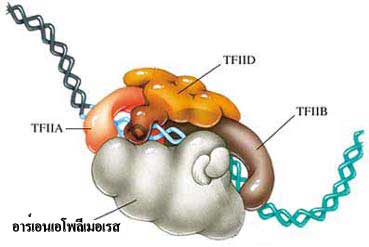
ภาพที่ 3.13 การทำงานของทรานสคริปชันแฟกเตอร์ (TF)ในกระบวนการคัดลอกดีเอนเอ
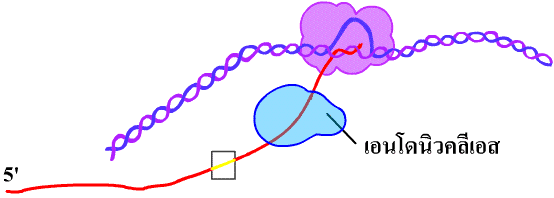
Elongation เป็นขั้นต่อสาย RNA ให้ยาวขึ้นโดยมีอีลองเกชันแฟกเตอร์(elongation factor)ช่วยในการสร้าง โดยอาร์เอนเอโพลีเมอเรสทำหน้าที่นำนิวคลีโอไทด์เข้ามาต่อสาย RNA ในทิศทาง 5'-3' บริเวณปลาย 5' (ส่วนต้น) ของ RNA ที่กำลังสร้างจะแยกออกจาก DNA และปล่อยให้ DNA 2 สายพันกันเป็นเกลียวดังเดิม โดยจะมีเฉพาะช่วงปลาย 3' ช่วงสั้นๆที่กำลังเติมนิวคลีโอไทด์ที่ยังพันอยู่กับ DNA (ภาพที่ 3.14) และเนื่องจากยีนต่างๆที่ใช้เป็นแม่แบบในการคัดลอกมีตำแหน่งกระจายตัวอยู่บน DNA ทั้ง 2 สาย ดังนั้นการสังเคราะห์ RNA สามารถเกิดขึ้นพร้อมกันได้ในหลายๆยีนบน DNA โมเลกุลเดียวกัน
![]()
![]()
![]()
ภาพที่ 3.14 ขั้นตอนต่อสาย (อีลองเกชัน) ของการคัดลอก
Termination เป็นขั้นตอนหยุดการสังเคราะห์ RNA โดยอาศัยโครงสร้างร่วมกับเทอร์มิเนเตอร์ (terminator) หรือ รหัสหยุด (termination signal) บน DNA ที่ประกอบด้วยเบส A ต่อกันเป็นจำนวนมากเมื่อ RNA สร้างมาถึงตำแหน่งนี้ อาร์เอนเอโพลีเมอเรสจะไม่สามารถนำนิวคลีโอไทด์มาจับกับ DNA แม่แบบได้อีก จึงหยุดการสร้างสาย RNA และจัดโครงสร้าง RNA เป็นรูปห่วง (hairpin loop) (ภาพที่ 3.15)
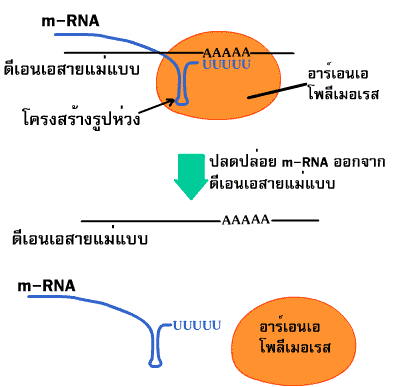
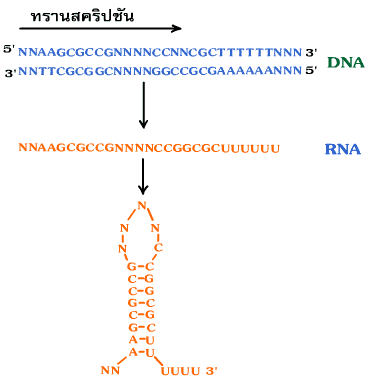
ภาพที่ 3.15 ขั้นตอนหยุด (เทอร์มิเนชัน) ของการคัดลอก
![]()
คำถาม : |
"ในการคัดลอก
DNA มาเป็น RNA นั้น มี DNA เพียงสายเดียว (จาก 2 สาย) ที่สังเคราะห์
RNA ได้" ท่านคิดเห็นอย่างไรกับข้อความนี้ |


หลังจากสังเคราะห์ได้ RNA ขึ้นมาแล้ว จะมีการตัดแต่งให้สมบูรณ์ โดยใน mRNA จะมีการเติม 7-methyl guanosine triphosphate เข้าไปที่ปลาย 5' (capping) (ภาพที่ 3.16) โดยใช้เอนไซม์กัวนีลิลทรานสเฟอเรส (guanylyl transferase) และเติมนิวคลีโอไทด์ที่มีเบสอะดินีน (A) หลายโมเลกุลเข้าที่ปลาย 3' ที่เรียกว่าโพลีอะดีนิเลชัน (polyadenylation) (ภาพที่ 3.17) รวมทั้งตัดส่วนอินทรอนทิ้งเพื่อให้ได้ mRNA ที่สมบูรณ์ (intron splicing) (ภาพที่ 3.18) จากนั้นจึงนำ RNA ที่สังเคราะห์ได้ส่งออกนอกนิวเคลียสไปสู่ไซโตพลาสซึม
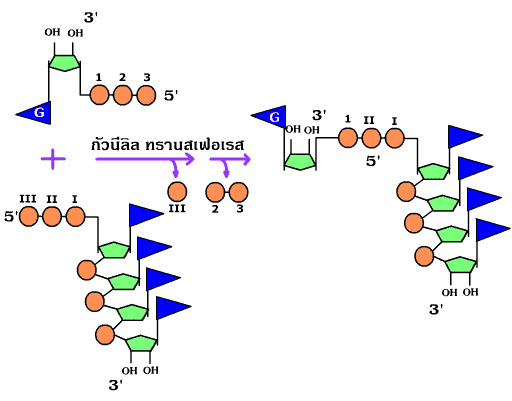
ภาพที่ 3.16 การเติม 7-methyl guanosine triphosphate เข้าที่ปลาย 5' ของ mRNA
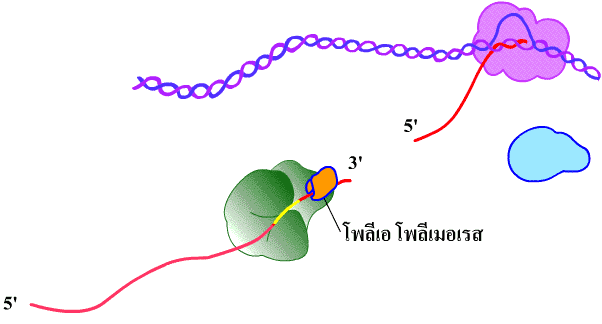

ภาพที่ 3.17
ขั้นตอนเติมนิวคลีโอไทด์ที่มีเบสอะดีนีนหลายโมเลกุล (โพลีอะดีนิเลชัน) ที่ปลาย
3' ของสาย mRNA
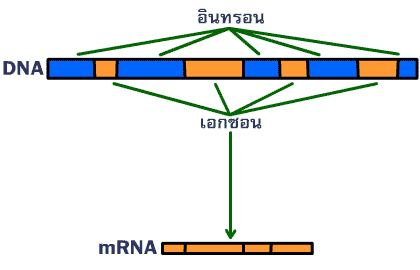
ภาพที่ 3.18 การตัดอินทรอนในสาย mRNA ที่สร้างใหม่ออกเพื่อให้ได้ mRNA ที่สมบูรณ์
![]()
คำถาม : |
ขบวนการตัดแต่ง
mRNA ในโปรคาริโอตและยูคาริโอต ต่างกันหรือไม่ อย่างไร |
| ภาพยนตร์ที่เกี่ยวข้องกับบทเรียน |
|
ชื่อเรื่อง |
ขนาด |
| transcription movie | 11,145 KB |
| mRNA modification movie | 10,808 KB |

