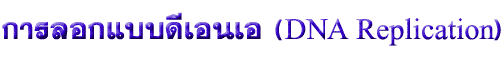|
|
||||||||
|
ยีนซึ่งเป็นดีเอนเอเกลียวคู่นั้น ก่อนเกิดการลอกแบบเพื่อสร้าง DNA สายใหม่ หรือคัดลอกเพื่อให้เป็น RNA จะต้องมีการคลายเกลียวออก การคลายเกลียวของ DNA ในนิวเคลียสซึ่งขดกันอยู่อย่างแออัดยัดเยียดนี้เป็นการสร้างปัญหาให้แก่สิ่งมีชีวิต จึงต้องมีกลไกการคลายเกลียวและคืนเกลียวอย่างพิเศษ คือ
|
||||||||
|
|
||||||||
ก่อนจะพูดถึงการแสดงออกของยีนซึ่งก็คือการคัดลอก และแปลรหัส จะพูดถึงเรื่องที่เกี่ยวข้อง คือการลอกแบบ DNA การลอกแบบ DNA ในสิ่งมีชีวิตทุกชนิดพบว่ามีลักษณะเฉพาะที่เหมือนกันหมดคือใช้สายเก่าเป็นแม่แบบในการสร้างสายใหม่ทุกครั้ง ดังนั้น DNA ใหม่ที่ได้ 2 สายจึงประกอบด้วยโพลีนิวคลีโอไทด์สายเก่า 1 สายและสายใหม่ 1 สาย เรียกการลอกแบบลักษณะนี้ว่าการลอกแบบกึ่งอนุรักษ์ (semiconservative replication) (ภาพที่ 3.1) |
||||||||
|
||||||||
ภาพที่ 3.1 การลอกแบบ DNA แบบกึ่งอนุรักษ์
|
||||||||
|
||||||||
การลอกแบบ DNA เป็นการเพิ่มปริมาณสารพันธุกรรมที่เหมือนกันไว้ใช้ในการทำกิจกรรมต่างๆ ของเซลล์ และเพื่อถ่ายทอดไปสู่ลูกหลานต่อไปในกระบวนการแบ่งเซลล์ การลอกแบบของ DNA มีขั้นตอนต่างๆ ดังนี้ |
||||||||
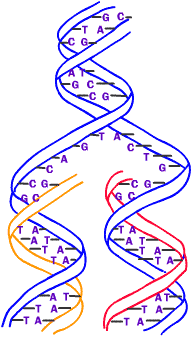
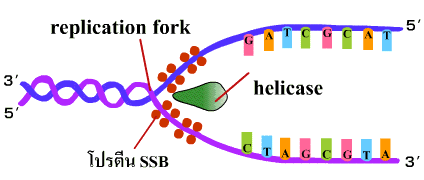
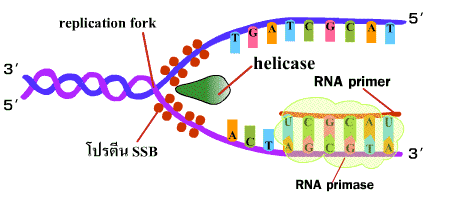
1. เอนไซม์เฮลิเคส (helicase) เข้าสลายพันธะไฮโดรเจนที่จุดใดจุดหนึ่งบนเกลียวคู่ (ภาพที่ 3.2) เรียกจุดนี้ว่าทางแยกของการลอกแบบหรือ เรพลิเคชันฟอร์ค (replication fork) |
||||||||
|
||||||||
ภาพที่ 3.2 เอนไซม์เฮลิเคสเริ่มสลายพันธะไฮโดรเจน |
||||||||
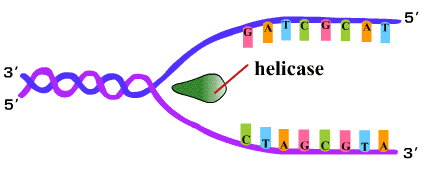
2. โปรตีน SSB (Single Strand Binding protein) เข้ามาเกาะบริเวณสายเดี่ยวที่แยกออกเพื่อป้องกันการพันเกลียวกลับ (ภาพที่ 3.3) |
||||||||
|
||||||||
ภาพที่ 3.3 โปรตีน SSB เข้าเกาะสาย DNA ที่แยกออก |
||||||||
3. เกิดการสร้าง RNA ชิ้นเล็กๆที่เรียกว่า อาร์เอนเอไพรเมอร์ (RNA primer) โดย อาร์เอนเอไพรเมส (RNA primase) ซึ่งไพรเมอร์นี้จะเป็นจุดที่เริ่มต้นการสังเคราะห์ DNA สายใหม่ (ภาพที่ 3.4) |
||||||||
|
|
||||||||
ภาพที่ 3.4 การสร้างอาร์เอนเอไพรเมอร์
|
||||||||
|
||||||||
|
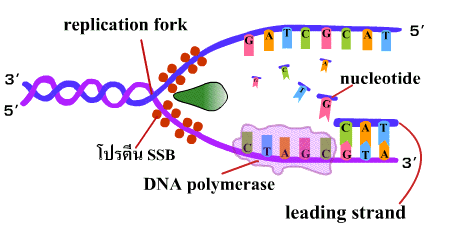
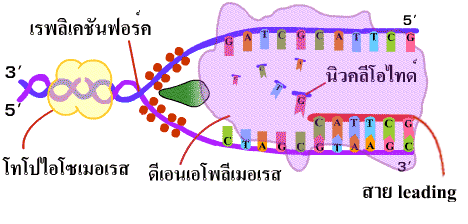
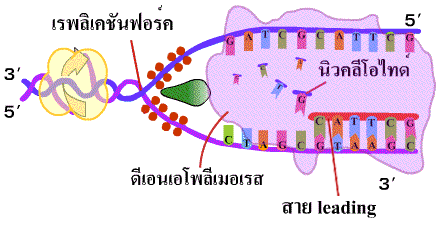
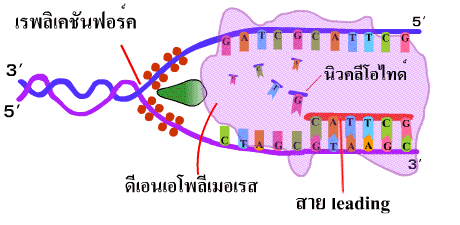
4. เอนไซม์ดีเอนเอโพลีเมอเรสนำดีออกซีไรโบนิวคลีโอไทด์เดี่ยว (deoxyribonucleotide) เข้ามาต่อสายในทิศทาง 5'-3' โดยเชื่อมเบสที่คู่กันเข้าด้วยกันด้วยพันธะไฮโดรเจน และเชื่อมหมู่ฟอสเฟตของแต่ละนิวคลีโอไทด์ให้เป็นพันธะฟอสโฟไดเอสเทอร์ เมื่อจุดคลายเกลียวเลื่อนขึ้นไปก็จะได้ DNA สายใหม่ที่ยาวขึ้น เรียก DNA สายนี้ว่า สายนำ (leading strand) (ภาพที่ 3.5) |
||||||||
|
|
||||||||
| ภาพที่ 3.5 การสร้าง DNA สาย leading |
||||||||
|
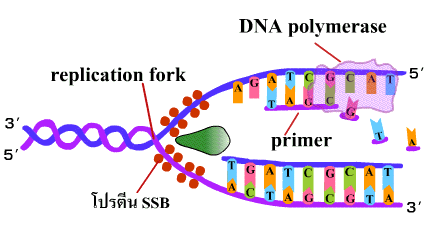
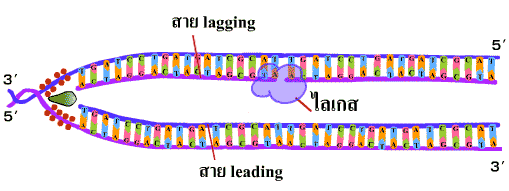
5. ในขณะที่เกิดการสังเคราะห์ DNA ขึ้นในสาย leading ก็จะเกิดการสังเคราะห์ DNA ในอีกสายหนึ่งซึ่งอยู่ตรงกันข้ามควบคู่กันไปด้วย สำหรับการสังเคราะห์ DNA ในสายตรงกันข้ามนี้เนื่องจากทิศทางการสังเคราะห์เป็นแบบ 5'-3' เสมอ เมื่อจุดคลายเกลียวเลื่อนขึ้น ไพรเมสจะสร้างอาร์เอนเอไพรเมอร์จับกับ DNA แม่แบบอีกสายหนึ่ง จากนั้นดีเอนเอโพลีเมอเรสจึงนำดีออกซีไรโบนิวคลีโอไทด์เข้ามาต่อเป็นสายไปทางด้านจุดเริ่มต้นการสร้าง (origin of replication) การทำเช่นนี้เป็นช่วงๆ จะได้เป็นชิ้น DNA สั้นๆ ที่ติดกับอาร์เอนเอไพรเมอร์ เรียกว่า ชิ้นส่วนโอกาซากิ (Okazaki fragment) อยู่ห่างกันเป็นช่วงๆ เรียก DNA สายนี้ว่า สายตาม(lagging strand) (ภาพที่ 3.6) |
||||||||
|
|
||||||||
|
|
||||||||
ภาพที่ 3.6 การสร้าง DNA สาย lagging |
||||||||
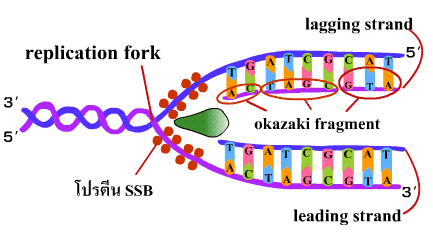
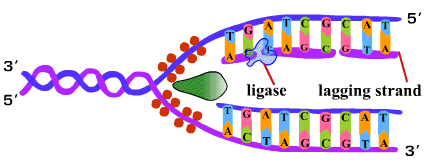
6. เมื่อการสังเคราะห์ DNA ดำเนินต่อไป ดีเอนเอโพลีเมอเรสจะมีการกำจัดอาร์เอนเอไพรเมอร์ออกด้วยคุณสมบัติของ 5'-3' เอ็กโซนิวคลีเอส (5'-3' exonuclease) และเติมดีออกซีไรโบนิวคลีโอไทด์ด้วยคุณสมบัติ 5'-3' โพลีเมอเรส และตามด้วยการเชื่อมพันธะฟอสโฟไดเอสเทอร์ของชิ้นส่วนโอกาซากิแต่ละโมเลกุลเข้าด้วยกันด้วยเอนไซม์ไลเกส (ligase) ในที่สุดได้เป็น DNA ใหม่ 2 โมเลกุล โดยแต่ละโมเลกุลมีสายเดิมอยู่ 1 สาย (ภาพที่ 3.7) |
||||||||
|
| ||||||||
|
| ||||||||
|
| ||||||||
ภาพที่ 3.7 การเชื่อม DNA สาย lagging
|
||||||||
|
||||||||
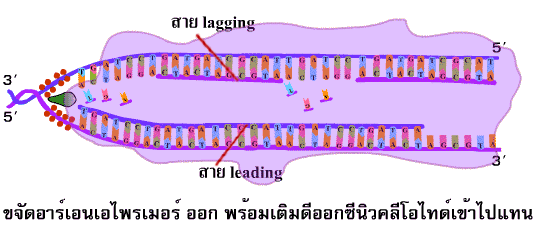
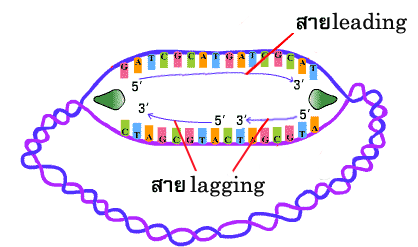
การสร้าง สาย leading และ สาย lagging สายใหม่มีทิศทางการสร้างที่สวนทางกัน (สาย leading สร้างยาวขึ้นในทิศทางการขยายของ ทางแยกของการลอกแบบ ส่วนสาย lagging สร้างยาวขึ้นไปทางด้านจุดเริ่มต้นของการสร้าง ซึ่งตรงข้ามกับการขยายของทางแยกของการลอกแบบ) เชื่อว่าการสร้างสายใหม่ทั้ง 2 สายนี้เกิดขึ้นพร้อมกัน (ภาพที่ 3.8 ) | ||||||||
| ||||||||
ภาพที่ 3.8 การทำงานของดีเอนเอโพลีเมอเรส ในการสร้างสาย lagging และ leading
|
||||||||
|
||||||||
การคลายเกลียวคู่ของ DNA ออกจากกันทำให้ส่วนเหนือขึ้นไปขดเป็นเกลียวแน่นขึ้น ซึ่งสามารถแก้ไขได้โดยเอนไซม์ โทโปไอโซเมอเรสซึ่งจะตัด DNA สายใดสายหนึ่งเหนือทางแยกของการลอกแบบ เพื่อให้สามารถคลายเกลียวที่แน่นออกได้และเชื่อมต่อใหม่ (ภาพที่ 3.9) | ||||||||
|
| ||||||||
|
| ||||||||
|
| ||||||||
ภาพที่
3.9 การทำงานของโทโปไอโซเมอเรส |
||||||||
|
|
||||||||
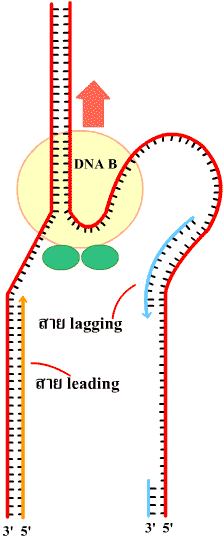
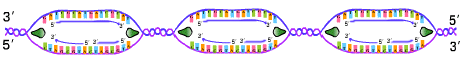
| ในการลอกแบบ DNA การขยายของทางแยกของการลอกแบบ (เรพลิเคชันฟอร์ค) จะเป็นแบบ 2 ทิศทางพร้อมกัน (bidirectional DNA replication) ทั้งในโปรคาริโอตและยูคาริโอต (ภาพที่ 3.10) |
||||||||
|
||||||||
|
||||||||
(บน) ในโปรคาริโอตเกิดทีละตำแหน่งในโครโมโซมวงแหวน (ล่าง)
ในยูคาริโอตเกิดพร้อมกันหลายตำแหน่งในโครโมโซมแท่ง
|
||||||||
|
||||||||
|
|
||||||||