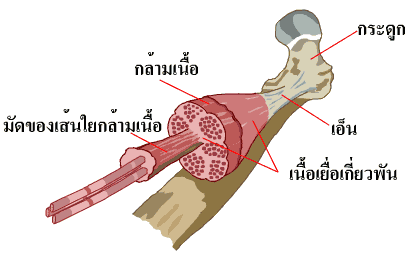
กล้ามเนื้อลายซึ่งห่อหุ้มร่างกายของเราไว้นั้นมีลักษณะเป็นมัดๆ มัดกล้ามเนื้อมีแผ่นเนื้อเยื่อเกี่ยวพัน (connective tissue) ซึ่งประกอบด้วยเส้นใยคอลลาเจน (collagen) และอีลาสติน (elastin) เป็นส่วนใหญ่หุ้มอยู่โดยรอบ ปลายข้างใดข้างหนึ่งหรือทั้งสองข้างของมัดกล้ามเนื้อเป็นคอลลาเจนที่ยึดติดกันเป็นเอ็น (tendon) ที่มีความเหนียวมากและยึดติดอยู่กับกระดูกอีกทีหนึ่ง เอ็นที่ปลายของกล้ามเนื้อบางชนิดอาจแผ่ออกเป็นแผ่นบางๆ (aponeuroses) ยึดติดกับกระดูกได้เป็นบริเวณกว้าง ภายในตัวกล้ามเนื้อมีเนื้อเยื่อเกี่ยวพันซึ่งเชื่อมต่อกับแผ่นด้านนอกแทรกอยู่โดยตลอด ใยประสาทและเส้นเลือดที่มาหล่อเลี้ยงกล้ามเนื้อจะซุกตัวอยู่ในเนื้อเยื่อเกี่ยวพันเหล่านี้
ี้
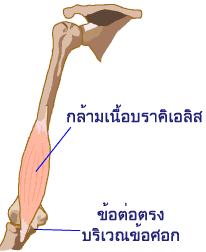
กล้ามเนื้อลายแต่ละมัดใช้ส่วนปลายที่เป็นเอ็นยึดกระดูกเอาไว้อย่างแน่นหนา เอ็นของมัดกล้ามเนื้อจะทอดตัวเป็นสะพานข้ามผ่านข้อต่อของกระดูกอย่างน้อยหนึ่งข้อหรือมากกว่านั้น แต่โดยทั่วไปแล้วจะพาดข้ามสองข้อ ดังนั้นเวลากล้ามเนื้อหดตัวจึงพาให้กระดูกหนึ่งชิ้น สองชิ้น หรือมากกว่าขยับเคลื่อน
เมื่อกล้ามเนื้อหดตัว ปลายเหนียวที่เป็นเอ็นจะยืดออกคล้ายๆ กับการยืดตัวของสปริง ในขณะที่เอ็นถูกยืดจะเกิดแรงตึงที่ไปดึงให้กระดูกเคลื่อนที่ การเคลื่อนที่ไปตามทิศทางที่ต้องการต้องอาศัยการทำงานของกล้ามเนื้อกลุ่มนำร่องที่เรียกว่า prime movers และยังต้องอาศัยการทำงานของกล้ามเนื้อชุดที่ทำงานเสริม (synergistic muscles) ที่จะทำให้ทิศทางการเคลื่อนที่ถูกต้องยิ่งขึ้น แถมยังช่วยทำหน้าที่ตรึง เพื่อไม่ให้ข้อต่อที่ไม่ต้องการเคลื่อนที่อีกด้วย
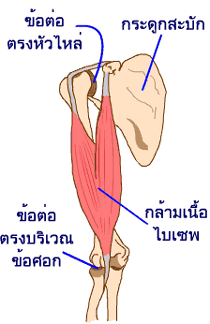
ในการเคลื่อนที่ของกระดูกกลับที่เดิม กล้ามเนื้อชุดที่พามันเคลื่อนที่มาไม่สามารถออกแรงดันได้เอง มันจึงต้องมีทีมกล้ามเนื้อคู่หู (antagonistic muscles) ที่ทำงานในสภาวะตรงกันข้าม มาช่วยทำงาน ทีมกล้ามเนื้อคู่หูจะทำหน้าที่ในการหดตัวเพื่อดึงกระดูกกลับมาที่ตำแหน่งเดิมหลังจากที่กล้ามเนื้อชุดแรกหดตัวแล้วพากระดูกให้เคลื่อนที่ไป
การทำงานของกล้ามเนื้อเป็นแรงดึงอย่างเดียว ไม่เป็นแรงผลัก
การงอแขนขึ้น เหยียดแขนลง เป็นตัวอย่างการทำงานร่วมกันของกล้ามเนื้อคู่หู
ในขณะที่กล้ามเนื้อชุดหนึ่งหดตัวเพื่อดึงกระดูกให้ขยับเคลื่อนไปด้านหนึ่งนั้น ทีมกล้ามเนื้อคู่หูก็จะคลายตัว เพื่อให้กล้ามเนื้อชุดแรกเคลื่อนที่รอบแกนของข้อต่อได้ตามความเหมาะสม แต่ก็ไม่ปล่อยเสียทีเดียว มันยังคงมีการหดตัวแต่เพียงน้อยๆ เพื่อให้การเคลื่อนไหวของกระดูกเป็นไปอย่างนุ่มนวล
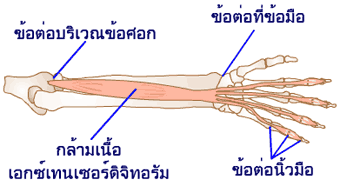
ผลการทำงานของกล้ามเนื้อลายทั้งหมดแบ่งออกได้เป็นสองประเภทคือ การงอ (flexion) และการเหยียด (extension) กล้ามเนื้อที่หดตัวแล้วทำให้อวัยวะนั้นงอเข้ามาเราเรียกว่า กล้ามเนื้อเฟลกเซอร์ (flexor) และกล้ามเนื้อที่หดตัวแล้วทำให้อวัยวะนั้นเหยียดออกเราเรียกว่า กล้ามเนื้อเอกซ์เทนเซอร์ (extensor)
การเคลื่อนไหวในบางลักษณะไม่สามารถจัดเข้าเป็นการงอ หรือการเหยียดได้อย่างชัดแจ้ง แต่เราพอจะจัดการเคลื่อนไหวต่างๆ ให้เข้าลักษณะสองประเภทนั้นได้ดังนี้
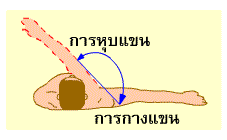
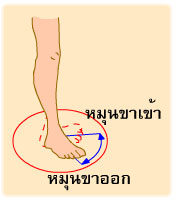
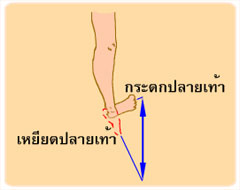
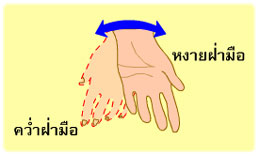
การงอ รวมถึง การหุบแขน หุบขา (adduction) การหมุนแขน หมุนขาเข้าหาลำตัว (medial rotation) การเหยียดปลายเท้าลง (plantar flexion of foot) การคว่ำฝ่ามือลง (pronation) การเหยียด รวมถึง การกางแขน กางขา (abduction) การหมุนแขน หมุนขาออกด้านนอก (lateral rotation) การกระดกปลายเท้าขึ้น (dorsi flexion of foot) และการหงายฝ่ามือขึ้น (supination)
การงอมักทำเพื่อให้เกิดงานต่างๆ ส่วนการเหยียดมักจะกระทำเพื่อคงท่าทางของร่างกาย
์
กล้ามเนื้อลายแต่ละมัดประกอบขึ้นด้วยเส้นใยกล้ามเนื้อ (muscle fiber) เรียงขนานกันเป็นจำนวนมาก เส้นใยกล้ามเนื้อแต่ละเส้นคือเซลล์หนึ่งเซลล์ เซลล์กล้ามเนื้อลายมีลักษณะเซลล์เป็นรูปทรงกระบอก มีความยาวตั้งแต่ 2-3 มิลลิเมตร ไปจนถึง 40 มิลลิเมตร และมีเส้นผ่าศูนย์กลางขนาด 10 ไมโครเมตร ถึง 100 ไมโครเมตร เมื่อเปรียบเทียบกับเซลล์อื่นในร่างกาย เราจะพบว่าเซลล์กล้ามเนื้อลายมีขนาดใหญ่กว่าเซลล์ร่างกายโดยทั่วไปที่มีเส้นผ่าศูนย์กลางประมาณ 10 ไมโครเมตร เท่านั้นเอง
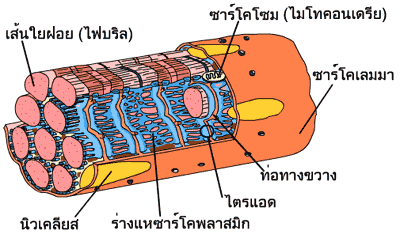
เซลล์กล้ามเนื้อแต่ละเซลล์มีนิวเคลียส (nucleus) อยู่เป็นจำนวนมาก นั่นเป็นเพราะว่าเซลล์กล้ามเนื้อเกิดจากการรวมตัวของเซลล์กล้ามเนื้อตั้งต้น (myoblast) นับร้อยเซลล์ตั้งแต่ตอนที่เรายังเป็นตัวอ่อนอยู่ในท้องแม่ นิวเคลียสของเซลล์กล้ามเนื้อจะอยู่ชิดกับเยื่อหุ้มเซลล์เลยทีเดียว องค์ประกอบต่างๆ ภายในเซลล์ก็มีเช่นเดียวกับเซลล์โดยทั่วไป แต่ชื่อที่เรียกองค์ประกอบนั้นๆ อาจจะแตกต่างออกไป เยื่อหุ้มเซลล์กล้ามเนื้อ เราเรียกว่า ซาร์โคเลมมา (sarcolemma) ส่วนของเหลวที่อยู่ภายในเซลล์เรียกว่า ซาร์โคพลาซึม (sarcoplasm) ซึ่งจะมีมัดของเส้นใยฝอย ที่มีชื่อเรียกในวงการกล้ามเนื้อว่าไฟบริล (fibril) ลอยตัวอยู่เกือบเต็มพื้นที่ของเซลล์ ช่องว่างระหว่างเส้นใยฝอยจะมีไมโทคอนเดรีย (mitochondria) ซึ่งในเซลล์กล้ามเนื้อเรียกว่าซาร์โคโซม (sarcosome) และร่างแหเอนโดพลาสมิก (endoplasmic reticulum) ที่มีชื่อเรียกว่า ร่างแหซาร์โคพลาสมิก (sarcoplasmic reticulum) แทรกตัวอยู่โดยตลอด
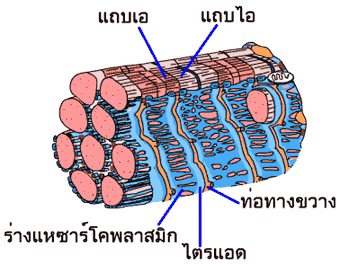
ร่างแหซาร์โคพลาสมิกของกล้ามเนื้อลายมีลักษณะเป็นท่อที่เชื่อมโยงกันโอบล้อมเส้นใยฝอยแต่ละเส้นไว้โดยรอบตามความยาว บางตอนของร่างแหซาร์โคพลาสมิกจะมีลักษณะแผ่บานออก ภายในส่วนบานนี้จะบรรจุแคลเซียมไอออน (Ca2+) ไว้มากมาย แคลเซียมไอออนพวกนี้จะจับอยู่กับโปรตีนชื่อแคลซีเควสตริน (calsequestrin) เส้นใยฝอยยังถูกล้อมรอบด้วยท่อทางขวาง (T-tubule หรือ transverse tubule) อีกด้วย ท่อทางขวางเป็นส่วนหนึ่งของเยื่อหุ้มเซลล์ที่ยื่นเข้าไปในเซลล์
ท่อทางขวางทำหน้าที่เป็นท่อส่งสัญญาณจากภายนอกเข้าไปยังเส้นใยกล้ามเนื้อทุกๆ เส้น ในบางบริเวณท่อทางขวางจะถูกขนาบด้วยส่วนแผ่บานของร่างแหซาร์โคพลาสมิก บริเวณดังกล่าวนั้นเรียกว่าไตรแอด (triad) ลักษณะเช่นนี้มีความสำคัญต่อกระบวนการการหดตัวของกล้ามเนื้อ
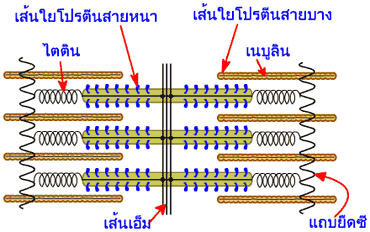
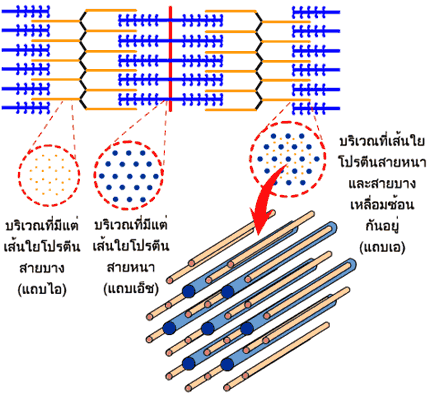
เซลล์กล้ามเนื้อหรือเส้นใยกล้ามเนื้อแต่ละเส้นประกอบขึ้นด้วยมัดของเส้นใยฝอยที่เรียกว่า ไฟบริล (fibril) ลอยตัวอยู่ในซาร์โคพลาสซึมจนเกือบเต็มพื้นที่ เส้นใยฝอยมีเส้นผ่าศูนย์กลางประมาณ 1 ไมโครเมตร เรียงขนานกันอยู่ตามความยาวของเส้นใยกล้ามเนื้อ เส้นใยฝอยแต่ละเส้นยังประกอบขึ้นด้วยหน่วยย่อยลงไปอีกเรียกว่า เส้นใยขนาดเล็ก (filament) เส้นใยขนาดเล็กมีอยู่สองชนิด คือ ชนิดที่เป็นเส้นใยโปรตีนสายหนา (thick filament) และชนิดที่เป็นเส้นใยโปรตีนสายบาง (thin filament)
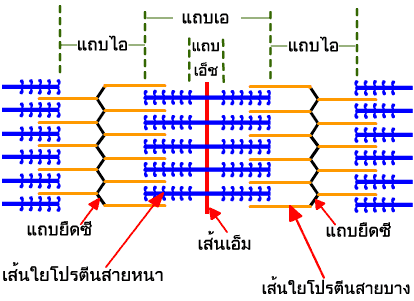
การเรียงตัวของเส้นใยโปรตีนสายหนาและเส้นใยโปรตีนสายบางทำให้เกิดเป็นหน่วยปฏิบัติการเพื่อการหดตัวของเส้นใยกล้ามเนื้อ (sarcomere) ซึ่งเป็นหน่วยย่อยที่สุดของมัดกล้ามเนื้อที่มีการหดตัวได้ เส้นใยฝอยแต่ละเส้นจะมีหน่วยปฏิบัติการย่อยดังกล่าวนี้เป็นจำนวนมากเรียงต่อกันไปตลอดความยาว แต่ละหน่วยปฏิบัติการมีเส้นคั่นตามขวาง เส้นนี้มีชื่อว่า แถบยืดซี (Z-disc ออกเสียงว่า ซี-ดิสก์)
เส้นใยกล้ามเนื้อเมื่อนำมาย้อมสี แล้วนำไปส่องดูด้วยกล้องจุลทรรศน์ จะเห็นเป็นลายตามขวาง ลายเหล่านี้ปรากฏอยู่ตรงตำแหน่งเดียวกันในเส้นใยฝอยทุกเส้นที่มารวมตัวกันเป็นเส้นใยกล้ามเนื้อ ทำให้เซลล์ของกล้ามเนื้อลายมีลักษณะลายสมชื่อของมัน
ลายในเส้นใยฝอยเกิดจากการเรียงตัวของโปรตีนองค์ประกอบในหน่วยปฏิบัติการย่อยของเส้นใยกล้ามเนื้ออย่างมีระเบียบแบบแผน ทำให้เราเห็นเป็นแถบทึบและแถบจางสลับกันไปเป็นระยะๆ
ภาพที่ 2.7 ภาพตัดบางตามความยาวแสดงหน่วยปฏิบัติการย่อยของเส้นใยกล้ามเนื้อ ซึ่งเรียงต่อกันตามความยาวของเส้นใยฝอย
หน่วยปฏิบัติการของเส้นใยกล้ามเนื้อแต่ละหน่วยจะมีอาณาเขตเริ่มต้นจากแถบยืดซี (Z-disc) หนึ่งแถบไปสิ้นสุดตรงแถบยืดซีอันถัดไป แต่ละหน่วยมีความยาวประมาณ 2.3 ไมโครเมตร
แถบทึบที่ครอบครองพื้นที่ตรงกลางหน่วยปฏิบัติการย่อยของกล้ามเนื้อ มีชื่อเรียกในวงการว่า แถบเอ (A-band หรือ anisotropic band) เกิดจากการที่มีเส้นใยโปรตีนสายหนาซึ่งมีเส้นผ่าศูนย์กลางประมาณ 15 นาโนเมตร เรียงตัวอยู่ โดยที่ปลายทั้งสองข้างของเส้นใยโปรตีนสายหนามีการเหลื่อมซ้อนกับปลายด้านหนึ่งของเส้นใยโปรตีนสายบาง ส่วนแถบจางที่อยู่ชิดแถบยืดซีทั้งสองข้างมีชื่อเรียกว่า แถบไอ (I-band หรือ isotropic band) จะมีเส้นใยโปรตีนสายบางที่มีเส้นผ่าศูนย์กลางประมาณ 7 นาโนเมตร เรียงตัวอยู่ ทำให้มีความโปร่งแสงกว่าเส้นใยโปรตีนสายหนา
anisotropic อ่านว่า แอน-ไอ-โซ-โทร-ปิก แปลว่า ไม่สม่ำเสมอ
สังเกตจากภาพ anisotropic band ทำให้ความเข้มของภาพไม่สม่ำเสมอ
ส่วนความยาวหนึ่งนาโนเมตรเท่ากับเศษหนึ่งส่วนพันล้านเมตรน่ะ
แถบจางอีกแถบหนึ่งซึ่งแฝงตัวอยู่ตรงกลางแถบเอ เราเรียกว่า แถบเอ็ช (H-zone) จะเป็นบริเวณที่ไม่มีส่วนของเส้นใยโปรตีนสายบางยื่นเข้ามาเลย
ปลายของเส้นใยโปรตีนสายบางด้านหนึ่งจะยึดติดอยู่กับแถบยืดซี โดยมีโปรตีนแอลฟาแอกตินิน (-actinin) ช่วยยึดเอาไว้ ส่วนบริเวณตรงกลางของเส้นใยโปรตีนสายหนาจะถูกตรึงเอาไว้ตรงเส้นเอ็ม (M-line หรือ M-protein) ซึ่งตั้งอยู่ตรงกึ่งกลางของแถบเอ็ช เส้นใยโปรตีนสายหนาที่อยู่สองฟากฝั่งของเส้นเอ็มจะเรียงตัวในทิศทางที่ตรงข้ามกัน ดังนั้นเส้นเอ็มจึงเป็นเส้นแบ่งครึ่งหน่วยปฏิบัติการเพื่อการหดตัวของเส้นใยกล้ามเนื้อ
ในหน่วยปฏิบัติการย่อยของเส้นใยกล้ามเนื้อ (sarcomere) ยังมีโปรตีนใหญ่ยักษ์อีกสองชนิด คือ ไตติน (titin) และเนบูลิน (nebulin) ที่ทำหน้าที่ขึ้นรูปให้กับเส้นใยโปรตีนสายหนาและเส้นใยโปรตีนสายบาง แต่ละโมเลกุลของโปรตีนไตตินจะทอดยาวจากเส้นเอ็มไปจนถึงแถบยืดซี ช่วยยึดเส้นใยโปรตีนสายหนาเอาไว้ให้อยู่ในตำแหน่งที่ถูกต้องตรงกลางของหน่วยปฏิบัติการย่อยของเส้นใยกล้ามเนื้อ นอกจากนี้มันยังมีคุณสมบัติยืดหยุ่นได้ ซึ่งมีประโยชน์ในการช่วยป้องกันไม่ให้หน่วยปฏิบัติการย่อยของเส้นใยกล้ามเนื้อถูกดึงทึ้งออกจากกันในขณะที่กล้ามเนื้อถูกยืด ส่วนโปรตีนเนบูลินทำหน้าที่เสมือนไม้บรรทัดที่คอยจัดระเบียบเส้นใยโปรตีนสายบางให้มีขนาดความยาวที่เหมาะสม
ถ้าเราตัดแถบเอตามขวางเราจะพบว่า เส้นใยโปรตีนสายหนาแต่ละเส้นถูกล้อมรอบไปด้วยเส้นใยโปรตีนสายบาง 6 เส้นด้วยกัน เรียงตัวได้ระยะเป็นรูปหกเหลี่ยม
ภาพที่ 2.12 ภาพตัดขวางแสดงการจัดเรียงตัวของเส้นใยโปรตีน ในหน่วยปฏิบัติการย่อยของเส้นใยกล้ามเนื้อ
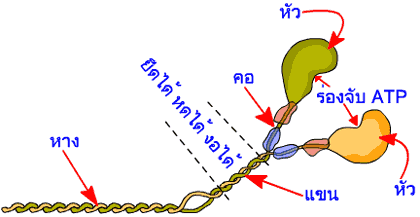
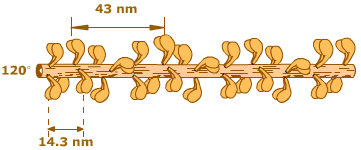
เส้นใยโปรตีนสายหนาของกล้ามเนื้อลายมีความยาว 1600 นาโนเมตร ประกอบขึ้นด้วยโปรตีนไมโอซิน (myosin) ประมาณ 300-400 หน่วย เรียงตัวซ้อนกันอยู่ โดยมีโปรตีนไตตินเป็นแกนกลาง แต่ละหน่วยของไมโอซินมีน้ำหนักโมเลกุลประมาณ 500 กิโลดาลตัน ประกอบด้วยเส้นใยโปรตีนขนาดใหญ่สองเส้น แต่ละเส้นมีน้ำหนักโมเลกุลประมาณ 200 กิโลดาลตัน พันกันเป็นเกลียวคู่ (double stranded helix) และมีความยาว 150 นาโนเมตร ปลายข้างหนึ่งของเส้นใยโปรตีนทั้งสองเส้นจะพับไปมาจนมีลักษณะมนๆ คล้ายหัวไม้กอล์ฟ เราเรียกส่วนนี้ว่าเป็นส่วนหัวของไมโอซิน จากส่วนหัวที่เรียกว่าคอยังมีโปรตีนขนาดเล็กอีกสองชนิด น้ำหนักโมเลกุลเส้นละประมาณ 20 กิโลดาลตัน เกาะอยู่ที่ส่วนคอข้างละคู่ ส่วนแขนของไมโอซิน อยู่ถัดมาจากส่วนคอ ส่วนก้านคอและส่วนปลายแขนจะเป็นสองบริเวณในโมเลกุลไมโอซินที่มีความยืดหยุ่น สามารถพับงอได้ บริเวณที่ถัดจากส่วนแขนเราเรียกเป็นส่วนหางของไมโอซิน
ภาพที่ 2.14 เส้นใยโปรตีนสายหนา
หัวของไมโอซินทั้งสองหัวนั้นจะมีบริเวณที่ให้ ATP มาเกาะอยู่ได้ เพราะส่วนหัวของไมโอซินมีคุณสมบัติเป็นเอนไซม์อะดีโนซีนไตรฟอสฟาเตส (myosin-adenosine triphosphatase หรือเรียกย่อๆ ว่า myosin-ATPase) ที่มีคุณสมบัติในการเร่งการสลายตัวของ ATP เป็น ADP และฟอสเฟตอนินทรีย์ (Pi) เพื่อให้ได้พลังงานสำหรับใช้ในการหดตัวของกล้ามเนื้อ
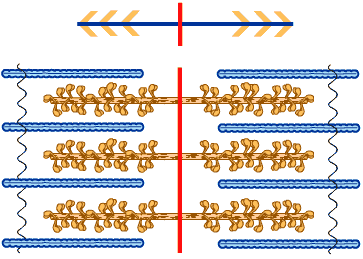
หน่วยปฏิบัติการย่อยของเส้นใยกล้ามเนื้อแต่ละหน่วยจะมีเส้นใยโปรตีนสายหนาหลายเส้นเรียงขนานกันอยู่ เส้นใยโปรตีนสายหนาที่อยู่สองฟากฝั่งของเส้นเอ็มจะเรียงตัวในทิศทางที่ตรงข้าม โดยใช้ส่วนหางของไมโอซินเชื่อมกันเอาไว้
โมเลกุลของไมโอซินที่อยู่รวมกันในเส้นใยโปรตีนสายหนาแต่ละเส้นจะยื่นส่วนหัว ส่วนคอ และส่วนแขน ออกไปเป็นกิ่ง ในทิศทางต่างๆ รอบตัว ในแต่ละรอบจะมีไมโอซินอยู่ 6 โมเลกุลด้วยกัน ดังนั้นเราจะพบหัวของไมโอซิน 12 หัวต่อหนึ่งรอบ หัวของไมโอซินที่ยื่นออกไปเป็นกิ่งนั้นมีความสามารถในการจับกับโปรตีนแอกทิน (actin) ของเส้นใยโปรตีนสายบางที่อยู่ล้อมรอบ การเกาะกันของแอกทินและไมโอซินทำให้เกิดเป็นสะพานเชื่อม (cross bridge) ระหว่างเส้นใยโปรตีนสายหนากับเส้นใยโปรตีนสายบางของเส้นใยกล้ามเนื้อ
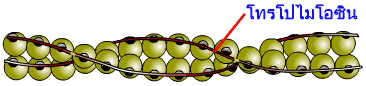
เส้นใยโปรตีนสายบางมีความยาว 1 ไมโครเมตร ประกอบด้วยโปรตีน 3 ชนิด คือ แอกทิน (actin) โทรโปไมโอซิน (tropomyosin) และโทรโปรนิน (troponin) โปรตีนแอกทินร่วมกับโปรตีนไมโอซินของเส้นใยโปรตีนสายหนาทำหน้าที่เกี่ยวกับการหดตัวและคลายตัวของกล้ามเนื้อ ส่วนโปรตีนโทรโปไมโอซิน และโทรโปนิน ทำหน้าที่ควบคุมกลไกดังกล่าว โปรตีนแอกทินเป็นโปรตีนที่มีลักษณะกลม (globular-actin เรียกย่อว่า G-actin) มีน้ำหนักโมเลกุล 43 กิโลดาลตัน แอกทินตัวกลม 375 หน่วย เรียงตัวต่อกันเป็นสาย (fibrous-actin หรือ F-actin) ดูคล้ายสายลูกประคำ สายแอกทินสองสายจะพันกันเป็นเกลียวเวียนขวา (right-handed double stranded helix) โดยมีระยะห่างของทุกช่วงเกลียวเท่ากันคือ ประกอบด้วยแอกทิน 7 หน่วย แต่ละหน่วยของโปรตีนแอกทินจะมีตำแหน่งสำหรับให้ส่วนหัวของไมโอซินมาเกาะอยู่ได้
โมเลกุลของโปรตีนที่ชื่อว่า โทรโปไมโอซิน ประกอบไปด้วยเส้นใยโปรตีนสองสาย แต่ละสายมีโครงสร้างเป็นเกลียว (-helical structure) โครงสร้างเกลียวสองสายนี้จะพันกันอีกทีเป็นซูเปอร์เกลียว (coiled-coil structure) ที่มีความแข็งแกร่งทนทานมาก แต่ละหน่วยของโทรโปไมโอซินมีความยาว 40 นาโนเมตร เรียงตัวต่อกันชนิดหัวจรดหางอยู่บนเกลียวของสายแอกทินเส้นคู่ แต่ละโมเลกุลของโทรโปไมโอซินมีความยาวเทียบเท่ากับช่วงเกลียวของแอกทินตัวกลม 7 หน่วย ที่อยู่บนสายแอกทิน
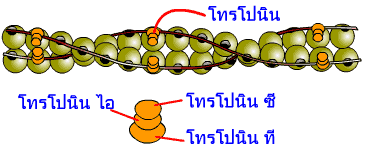
โปรตีนควบคุมที่ชื่อ โทรโปนิน เป็นโปรตีนที่มีขนาดเล็ก จับอยู่กับโทรโปไมโอซิน ทุกโมเลกุลของโทรโปนินประกอบด้วย 3 หน่วยย่อย คือ โทรโปนิน ที (troponin T) โทรโปนิน ซี (troponin C) และโทรโปนิน ไอ (troponin I) โทรโปนิน ที เป็นหน่วยย่อยที่ทำหน้าที่เป็นตัวยึดโมเลกุลของโทรโปนินเข้ากับโทรโปไมโอซิน โทรโปนิน ซี เป็นหน่วยที่มีความสามารถในการจับตัวกับแคลเซียมไอออน ซึ่งจะเป็นตัวเริ่มกระบวนการการหดตัวของกล้ามเนื้อ การจับกันของแคลเซียมไอออนกับโทรโปนิน ซี มีผลทำให้โทรโปนิน ซี มีการเปลี่ยนแปลงโครงร่างไป ซึ่งจะส่งผลต่อการเปลี่ยนแปลงรูปร่างของโทรโปนิน ที และ โทรโปนิน ไอ ด้วย โทรโปนิน ที ที่เปลี่ยนแปลงรูปร่างไปในขณะที่มีแคลเซียมมาเกาะกับโทรโปนิน ซี นั้นจะเป็นตัวดึงโปรตีนโทรโปไมโอซินให้เปลี่ยนตำแหน่งเกาะบนสายแอกทินเล็กน้อย ทำให้ตำแหน่งเกาะของไมโอซินบนสายแอกทินเปิดออก หัวของไมโอซินก็สามารถเกาะกับแอกทินได้ สำหรับโทรโปนิน ไอ ทำตัวเป็นคู่แข่งกับโทรโปนิน ที ในขณะที่ไม่มีแคลเซียมไอออนมาจับกับโทรโปนิน ซี มันจะเปลี่ยนแปลงรูปร่างไป ส่งผลให้โทรโปนิน ที ดึงโทรโปไมโอซินกลับไปอยู่ในตำแหน่งที่ขวางทางการเกาะของไมโอซินกับแอกทิน