|

|
กล้ามเนื้อของเรานับเป็นประดิษฐกรรมชั้นเยี่ยม
ที่มีกลไกการทำงานสลับซับซ้อน นักวิทยาศาสตร์พยายามหาคำอธิบายเกี่ยวกับกลไกการทำงานของกล้ามเนื้อโดยใช้หลักฐานที่พบจากการใช้กล้องจุลทรรศน์อิเล็กตรอน
ซึ่งมีกำลังขยายสูงกว่ากล้องจุลทรรศน์แบบธรรมดาถึงหนึ่งพันเท่า
จากการศึกษาที่ทำโดยการยืดกล้ามเนื้อออกและตรึงกล้ามเนื้อเอาไว้
แล้วทำการกระตุ้นให้กล้ามเนื้อเกิดการหดตัว นักวิทยาศาสตร์พบว่าในขณะที่กล้ามเนื้อเกิดการหดตัว
ความกว้างของแถบเอไม่ได้มีการเปลี่ยนแปลงเลย นอกจากนี้ความยาวของเส้นใยโปรตีนสายหนาและสายบางก็ยังคงเดิม
ทั้งๆ ที่ความกว้างของหน่วยปฏิบัติการเพื่อการหดตัวของกล้ามเนื้อ
แถบไอและแถบเอ็ช มีการเปลี่ยนแปลงขนาดคือหดสั้นลง
|
|
ภาพที่ 3.1 ภาพตัดบางตามความยาวแสดงหน่วยปฏิบัติการย่อยของ
เส้นใยกล้ามเนื้อขณะพักงานและขณะหดตัว
|

|
กลไกการหดตัวของกล้ามเนื้อสามารถอธิบายได้ด้วยทฤษฎีการเลื่อนตัวของเส้นใยโปรตีนขนาดเล็ก
(sliding filament model) ที่เสนอโดยนักวิทยาศาสตร์ชาวอังกฤษสองกลุ่ม
คือ กลุ่มของ ฮิว ฮักซ์ลีย์ (Hugh Huxley) และ จีน แฮนสัน (Jean
Hanson) กับกลุ่มของ แอนดรูว์ ฮักซ์ลีย์ (Andrew Huxley) และ
ราล์ฟ นีเดอร์เกอร์กี (Ralph Niedergerke) เมื่อปี พ.ศ. 2497
กล่าวคือ การหดตัวของกล้ามเนื้อเกิดจากการเลื่อนของเส้นใยโปรตีนสายบางที่อยู่สองข้างของหน่วยปฏิบัติการเพื่อการหดตัวของกล้ามเนื้อ
เข้าหาเส้นใยโปรตีนสายหนา มีผลทำให้การเหลื่อมซ้อนกันของเส้นใยโปรตีนสายบางและเส้นใยโปรตีนสายหนาเพิ่มขึ้น
ยิ่งมีการเหลื่อมซ้อนกันมากขึ้นเท่าไร ความกว้างของแถบไอ ซึ่งเป็นบริเวณที่มีแต่เส้นใยโปรตีนสายบางอย่างเดียว
และบริเวณแถบเอ็ชซึ่งมีแต่เส้นใยโปรตีนสายหนาอย่างเดียวก็จะยิ่งหดสั้นตามไปด้วย
|
|
การเลื่อนของเส้นใยโปรตีนสายบางเข้าหาเส้นใยโปรตีนสายหนา
เกิดขึ้นได้โดยที่ไมโอซินยื่นส่วนหัวมาเกาะกับบริเวณจับจำเพาะบนโมเลกุลของแอกทินในเส้นใยโปรตีนสายบาง
แล้วดึงสายแอกทินให้เลื่อนเข้ามาสู่ศูนย์กลางของหน่วยปฏิบัติการเพื่อการหดตัวของกล้ามเนื้อ
ส่วนหัวของไมโอซินที่ยื่นเป็นกิ่งออกมาจากเส้นใยโปรตีนสายหนาแต่ละเส้นนั้น
จะเกาะกับโมเลกุลของแอกทินบนเส้นใยโปรตีนสายบางที่อยู่ล้อมรอบทั้ง
6 สาย การเกาะกันของแอกทินกับไมโอซิน รวมทั้งแรงที่ใช้ในการดึงเส้นใยโปรตีนสายบางอาศัยพลังงานที่เกิดจากการสลายตัวของ
ATP
|
| |
|
การหดตัวของกล้ามเนื้อในแต่ละครั้งจะมีการเกาะกันของไมโอซินกับแอกทินและการดึงเกิดขึ้นหลายครั้งเป็นวงจรต่อเนื่องกันไป
โดยที่ไมโอซินจะเปลี่ยนตำแหน่งที่เกาะบนสายแอกทินไปเรื่อยๆ มุ่งหน้าไปทางแถบยืดซี
แต่ละโมเลกุลของไมโอซินที่ทอดตัวเป็นสะพานเชื่อมกับแอกทินนั้นประกอบด้วยส่วนหัวสองหัว
ไมโอซินแต่ละหัวซึ่งมีคุณสมบัติเป็นเอนไซม์ myosin-ATPase จะมีร่องที่ให้
ATP เข้ามาเกาะได้ 1 โมเลกุล
|
|
การเกาะกันของ
ATP กับไมโอซินจะมีผลทำให้แอกทินหลุดออกจากการเกาะกับไมโอซิน
บริเวณเร่งของเอนไซม์ myosin-ATPase จะปิดล้อม ATP เอาไว้ |
|
 |
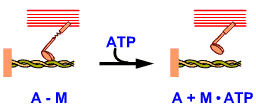
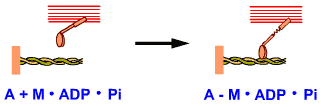
A-M
หมายถึง การเกาะกันระหว่างแอกทินกับไมโอซิน
A+M.ATP หมายถึง
สายแอกทิน กับ สายไมโอซินที่มี ATP เกาะอยู่และหลุดจากการเกาะกับแอกทิน |
|
เมื่อเอนไซม์ทำการสลาย
ATP แล้ว ผลิตผลที่เกิดขึ้นคือ ADP และฟอสเฟตอนินทรีย์ จะยังคงเกาะอยู่กับเอนไซม์
ส่วนพลังงานที่ถูกปลดปล่อยออกมาจะถูกส่งให้กับโปรตีนไมโอซิน
ทำให้ไมโอซินอยู่ในสภาวะที่มีพลังงานสูง มีการเปลี่ยนแปลงโครงร่างไปเป็นแบบที่มีความสามารถในการจับกับแอกทิน
นั่นคือส่วนหัวของมันจะทำมุมตั้งฉากกับส่วนหาง แล้วเข้าเกาะกับแอกทินอย่างรวดเร็ว
การจับกันระหว่างแอกทินกับไมโอซินทำให้เกิดเป็นโปรตีนยักษ์เชิงซ้อน
ที่เรียกว่า actomyosin complex |
|
|
A+M.ADP.Pi
หมายถึง สายแอกทิน กับ สายไมโอซินที่มี ADP และ
Pi ซึ่งได้จากการสลาย ATP เกาะอยู่
A-M.ATP หมายถึง การเกาะกันระหว่างแอกทินกับ
ไมโอซินโดยที่ ADP และ Pi ยังคงเกาะอยู่กับไมโอซิน |
|
เมื่อส่วนหัวของไมโอซินสลัดฟอสเฟตอนินทรีย์ออกไป
มันจะสามารถเกาะกับแอกทินได้กระชับขึ้น พลังงานที่สะสมเอาไว้จะทำให้โครงร่างของไมโอซินมีการเปลี่ยนแปลงอย่างมโหฬาร
ส่วนหัวที่เคยทำมุม 90 องศากับส่วนหางจะเปลี่ยนไปทำมุม 45 องศา
การเปลี่ยนแปลงโครงร่างของไมโอซินในขณะที่กำลังเกาะอยู่กับแอกทินอยู่นั้น
ทำให้เกิดความตึงบนเส้นใยโปรตีนสายบางที่มันเกาะอยู่ ทำให้เส้นใยโปรตีนสายบางถูกดึงให้เลื่อนผ่านเข้ามาสู่ศูนย์กลางของหน่วยปฏิบัติการเพื่อการหดตัวของกล้ามเนื้อ
วัดระยะทางได้ในราว 10 นาโนเมตร
|
|
|
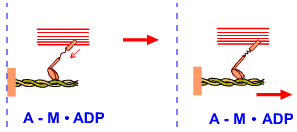
A-M.ADP หมายถึง
การเกาะกันระหว่างแอกทินกับไมโอซิน โดยที่ ADP ยังคงเกาะอยู่กับไมโอซิน |
|
ในขั้นตอนสุดท้าย
ADP ที่เกาะอยู่กับไมโอซินจะหลุดออก แต่ยังคงทิ้งให้ไมโอซินกับแอกทินเกาะเกี่ยวกันอยู่อย่างนั้น
จนกระทั่ง |
|
การเกาะกันของหัวไมโอซินกับแอกทินจะคงอยู่อย่างนั้นจนกระทั่งมี
ATP ใหม่เข้ามาเกาะตรงร่องจับบนไมโอซิน แล้วส่วนหัวของไมโอซินก็หลุดออกจากแอกทินในท่าทำมุม
45 องศา กับส่วนหาง และพร้อมที่จะกลับไปเริ่มต้นวัฏจักรการจับปล่อยอีกครั้ง
|
|
วัฏจักรการจับปล่อยก็เป็นเช่นนี้เอง |
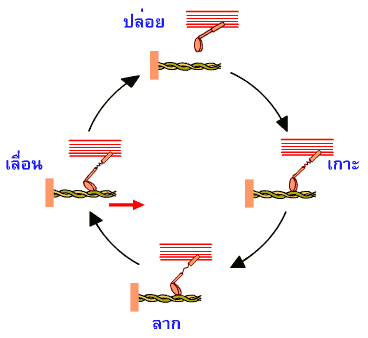
|
ภาพที่ 3.2 แสดงจังหวะการจับปล่อยของไมโอซินในวัฏจักรการจับปล่อยของแอกทินไมโอซิน
|
|
|
การเกาะกันระหว่างไมโอซินกับแอกทินในยกที่สอง
สามและยกต่อๆ มา ในขณะที่เกิดการหดตัวของกล้ามเนื้อ ส่วนหัวของไมโอซินจะไปจับกับแอกทินตรงตำแหน่งใหม่ที่อยู่ในทิศทางใกล้กับแถบยืดซีมากขึ้น
เพราะตำแหน่งที่เกาะเดิมได้เคลื่อนผ่านเส้นใยโปรตีนสายหนาไปแล้ว
ในแต่ละวินาทีส่วนหัวของไมโอซินจะเข้าสู่วังวนของการจับปล่อยได้ถึง
5 ครั้ง ติดต่อกัน |
|
ตัวเล็กอยากดูวงจรความสัมพันธ์ระหว่างแอกทิน
ไมโอซิน และ ATP ด้วยน่ะ พี่เนื้อลาย |
|
จะให้ดูอย่างละเอียดเลย
อย่าสับสนนะ |
|
|
|
ภาพที่ 3.3 แสดงวงจรความสัมพันธ์ระหว่างแอกทิน ไมโอซิน และ ATP
|
|
|
|
 |
การควบคุมการหดตัวและคลายตัวของกล้ามเนื้อเกิดขึ้นได้อย่างไรกันนี่ |
|
ทุกครั้งที่เราเคลื่อนไหว
สมอง ประสาท และกล้ามเนื้อ จะต้องทำงานสัมพันธ์กันเป็นอย่างดี
สมองรับข้อมูลจากประสาทสัมผัส ทำการแปลผล และตัดสินใจว่าจะตอบสนองให้ร่างกายมีการเคลื่อนไหวอย่างไร
สัญญาณประสาทจากระบบประสาทส่วนกลาง จากเซลล์ประสาทสั่งการ (motor
neuron-มอเตอร์นิวรอน) จะถูกส่งไปตามเส้นประสาทไขสันหลัง ไปยังปลายประสาทที่กล้ามเนื้อ
สัญญาณประสาทจะกระตุ้นให้กล้ามเนื้อทำงานโดยการหดตัว
|
|
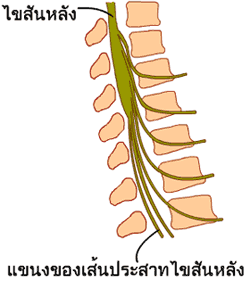
ภาพที่ 3.4 เส้นประสาทไขสันหลัง
|
กล้ามเนื้อลายแต่ละมัดมีเส้นประสาทมาหล่อเลี้ยงมากมาย
เส้นประสาทแต่ละเส้นที่มายังมัดกล้ามเนื้อจะมีการแตกแขนงออกไปเพื่อไปเลี้ยงเส้นใยกล้ามเนื้อได้เป็นจำนวนมาก
เส้นใยกล้ามเนื้อทั้งหมดที่ถูกหล่อเลี้ยงโดยแขนงประสาทที่มาจากเส้นประสาทไขสันหลังหนึ่งเส้น
จะประกอบขึ้นเป็นเส้นใยกล้ามเนื้อที่เป็นหน่วยทำงานหน่วยเดียวกัน
(motor unit) จำนวนเส้นใยกล้ามเนื้อของแต่ละหน่วยมีตั้งแต่หนึ่งเส้นไปจนกระทั่งถึงหลายร้อยเส้น
การมีเส้นใยกล้ามเนื้อมากหรือน้อยขึ้นอยู่กับหน้าที่และขนาดของกล้ามเนื้อนั้นๆ
กล้ามเนื้อที่ต้องการความละเอียดแม่นยำในการทำงาน เช่น กล้ามเนื้อที่ช่วยในการหยิบจับสิ่งของ
จำนวนเส้นใยกล้ามเนื้อที่ถูกควบคุมด้วยแขนงประสาทจากเส้นประสาทไขสันหลัง
1 เส้น จะน้อยกว่ากล้ามเนื้อที่ไม่ต้องทำงานด้วยความแม่นยำมากนัก
เช่น กล้ามเนื้อที่ทำหน้าที่เกี่ยวกับการทรงตัว |
|
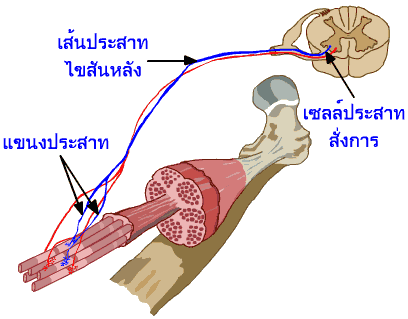
ภาพที่
3.5 แสดงแขนงประสาทของเส้นประสาทไขสันหลังที่มายังกล้ามเนื้อ |
|
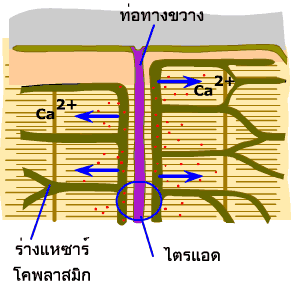
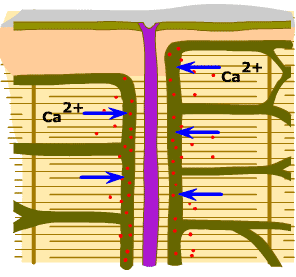
สัญญาณประสาทที่มายังกล้ามเนื้อ
จะถูกส่งผ่านไปตามเยื่อหุ้มเซลล์ และผนังด้านในของท่อทางขวาง
ไปสู่เส้นใยกล้ามเนื้อทุกๆ เส้นที่เส้นประสาทไขสันหลังนั้นควบคุมอยู่
ผนังของท่อทางขวางตรงส่วนที่ถูกขนาบด้วยส่วนแผ่บานของร่างแหซาร์โคพลาสมิก
จะมีช่องทางให้แคลเซียมไอออนจากภายนอกเซลล์ผ่านเข้ามา ช่องแคลเซียมไอออนนี้จะเปิดออกก็ต่อเมื่อมีสัญญาณไฟฟ้ามากระตุ้น
(voltage-gated Ca2+ channels) ทั้งนี้เนื่องจากโปรตีนที่ประกอบขึ้นเป็นช่องแคลเซียมไอออนเป็นโปรตีนที่มีส่วนที่ไวต่อความต่างศักย์
(voltage-sensitive) ช่องไอออนนี้ถูกยับยั้งได้ด้วยสารประกอบไดไฮโดรไพริดีน
(dihydropyridine) จึงมีชื่อเรียกว่าตัวรับไดไฮโดรไพริดีน
(dihydropyridine receptor) |
|
ภาพที่ 3.6 แสดงช่องแคลเซียมไอออนบนผนังท่อทางขวางและบนผนังร่างแหซาร์โคพลาสมิก
|
เมื่อสัญญาณประสาทของกล้ามเนื้อเคลื่อนมาตามผนังท่อทางขวาง
ช่องทางแคลเซียมไอออนจะเปิดออก แคลเซียมจากภายนอกเซลล์ที่หลั่งเข้ามาในท่อทางขวางจะข้ามไปยังช่องแคลเซียมไอออนของร่างแหซาร์โคพลาสมิกซึ่งเรียงตัวขนานกันอยู่
เพื่อส่งสัญญาณให้ร่างแหซาร์โคพลาสมิกหลั่งแคลเซียมไอออนออกมาในซาร์โคพลาซึมผ่านทางช่องแคลเซียมไอออนของมันเอง
ที่เรียกว่า ตัวรับไรอะโนดีน (ryanodine receptor) |
|
ภาพที่ 3.7 แสดงการหลั่งของแคลเซียมไอออนจากร่างแหซาร์โคพลาสมิกเข้าสู่ซาร์โคพลาซึม
|


|
|
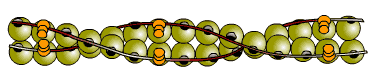
ในระยะพักทั้งเส้นใยโปรตีนสายหนาและสายบางต่างอยู่เป็นอิสระไม่มีกิ่งมาเชื่อมติดกัน
ทั้งนี้เนื่องจากมีการเปลี่ยนแปลงโครงร่างของโทรโปนินทั้งโมเลกุล
ที่ส่งผลทำให้โมเลกุลของโทรโปไมโอซินไปปิดบังตำแหน่งเกาะของไมโอซินบนแอกทินไว้
ทำให้ไมโอซินไม่สามารถเข้าเกาะกับแอกทินได้
|
|
|
ภาพที่ 3.8 โทรโปไมโอซินบังตำแหน่งเกาะของไมโอซินบนสายแอกทิน
|
แต่เมื่อกล้ามเนื้อถูกกระตุ้นด้วยสัญญาณประสาท
แคลเซียมไอออนที่หลั่งออกมาจากร่างแหซาร์โคพลาสมิกจะจับกับโทรโปนิน
ซี การรวมตัวของแคลเซียมไอออนกับโทรโปนิน ซี จะทำให้โทรโปนินทั้งโมเลกุลมีการเปลี่ยนแปลงรูปร่าง
ซึ่งจะมีผลต่อไปทำให้โมเลกุลของโทรโปไมโอซินเปลี่ยนตำแหน่งเกาะบนสายแอกทินไปเล็กน้อย
ตำแหน่งที่เกาะของไมโอซินบนแอกทินตัวกลมทั้ง 7 หน่วย ที่ถูกโทรโปไมโอซินบังไว้จะถูกเปิดออก |
|
ภาพที่ 3.9 ตำแหน่งเกาะของไมโอซินบนสายแอกทินเปิดออก
|
การเปิดออกของตำแหน่งเกาะของไมโอซินบนสายแอกทิน
ทำให้หัวของไมโอซินสามารถเข้าจับกับแอกทินได้ วัฏจักรการจับปล่อยจึงเกิดขึ้น |
|
|
|
โทรโปนิน
ซี สามารถจับแคลเซียมไอออนไว้ได้ชั่วหนึ่งรอบของวัฏจักรการจับปล่อยเท่านั้น
ทำให้กล้ามเนื้อไม่สามารถเข้าสู่วัฏจักรการจับปล่อยได้หลายๆ
รอบจนได้แรงตึงสูงๆ แคลเซียมไอออนจะถูกเก็บเข้าร่างแหซาร์โคพลาสมิกอย่างรวดเร็วโดยวิธีการปั๊ม
การปั๊มแคลเซียมไอออนกลับต้องอาศัยพลังงานจาก ATP โดยการทำงานของเอนไซม์
Ca2+ - ATPase ที่อยู่บริเวณผนังของร่างแหซาร์โคพลาสมิก
|
|
ภาพที่ 3.10 แคลเซียมไอออนจากซาร์โคพลาซึมถูกปั๊มกลับเข้าสู่ร่างแหซาร์โคพลาสมิก
|
|
การทำให้วัฏจักรการจับปล่อยสามารถเกิดวนไปวนมาจนกระทั่งกล้ามเนื้อหดตัวจนได้แรงตึงสูงสุดนั้น
ต้องทำให้แคลเซียมไอออนหลั่งออกมาในซาร์โคพลาซึมอย่างต่อเนื่อง
เหตุการณ์เช่นนี้จะเกิดขึ้นได้ต้องอาศัยการกระตุ้นซ้ำด้วยสัญญาณประสาทของกล้ามเนื้อ
|
 |
|
เมื่อกล้ามเนื้อหดตัว
หน่วยปฏิบัติการเพื่อการหดตัวของกล้ามเนื้อ มีปริมาตรคงที่
คุณคิดว่าเป็นเพราะเหตุใด |
|
การเคลื่อนของเส้นใยโปรตีนสายบางเข้าหาศูนย์กลางของหน่วยปฏิบัติการเพื่อการหดตัวของกล้ามเนื้อ
มีผลทำให้หน่วยปฏิบัติการฯ มีความยาวสั้นลง
|
|
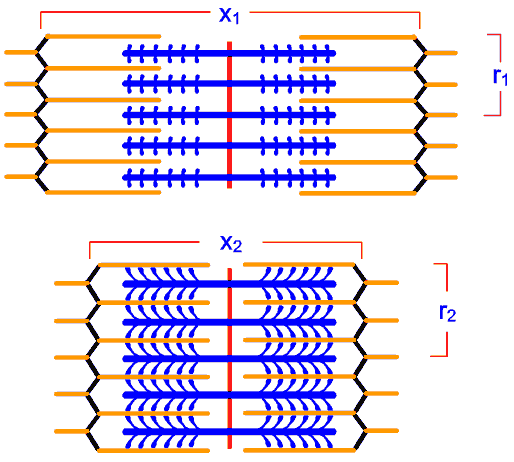
ภาพที่ 3.11 แสดงปริมาตรหน่วยปฏิบัติการย่อยของเส้นใยกล้ามเนื้อในขณะพัก
และขณะหดตัว
|
|
X1
แทน ความยาวของหน่วยปฏิบัติการย่อยของเส้นใยกล้ามเนื้อในขณะพัก
r1
แทน ครึ่งความสูงของหน่วยปฏิบัติการย่อยของเส้นใยกล้ามเนื้อในขณะพัก
X2
แทน ความยาวของหน่วยปฏิบัติการย่อยของเส้นใยกล้ามเนื้อในขณะหดตัว
r2
แทน ครึ่งความสูงของหน่วยปฏิบัติการย่อยของเส้นใยกล้ามเนื้อในขณะหดตัว |
|
|
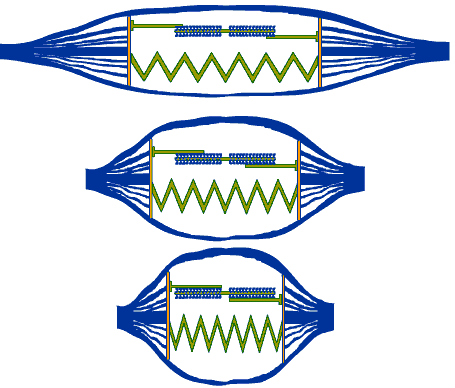
การทำงานของกล้ามเนื้ออาศัยความพร้อมเพรียงในการทำงานของหน่วยปฏิบัติการย่อยทุกๆ
หน่วย ที่ประกอบขึ้นเป็นเส้นใยฝอยของเส้นใยกล้ามเนื้อที่เป็นหน่วยทำงานเดียวกัน
ดังนั้นหากมีการหดตัวเกิดขึ้นพร้อมๆ กันทั้งหมด ก็จะทำให้กล้ามเนื้อทั้งมัดมีการหดตัวสั้นลงไปด้วย |
| |
ภาพที่ 3.12 แสดงความยาวของมัดกล้ามเนื้อที่มีการเปลี่ยนแปลงเมื่อกล้ามเนื้อหดตัว
|
|
|
คุณคิดว่าลูกหนูเกิดจากอะไร
ก. แม่หนู
ข. การหดตัวของกล้ามเนื้อ
|
|
|
|
เมื่อเราใช้งานกล้ามเนื้อจนพอใจแล้ว
ระบบประสาทก็จะสั่งการให้สัญญาณประสาทหยุดเดินทางมายังกล้ามเนื้อ
ทำให้แคลเซียมไอออนไม่ถูกหลั่งออกมาจากร่างแหซาร์โคพลาสมิกเข้าสู่ซาร์โคพลาซึม
ปฏิกิริยาการเกาะกันของแอกทินและไมโอซินจึงไม่สามารถเกิดขึ้นได้อีก
เส้นใยโปรตีนสายบางจะเคลื่อนกลับไปยังจุดเริ่มต้นผ่านเส้นใยโปรตีนสายหนา
หน่วยปฏิบัติการเพื่อการหดตัวของเส้นใยกล้ามเนื้อจะกลับมามีความยาวเท่ากับความยาวในระยะพัก
เมื่อทุกๆ หน่วยปฏิบัติการย่อยของเส้นใยกล้ามเนื้อเป็นอย่างนี้เหมือนกันหมด
เส้นใยกล้ามเนื้อรวมทั้งมัดกล้ามเนื้อก็จะคลายตัวกลับมามีความยาวเท่าเดิมด้วยเช่นกัน
|
|
|
เหตุใดคนที่เสียชีวิตไปสักค่อนวันแล้ว
จึงมีร่างกายที่แข็งเกร็ง
|
|
|

|
|
ตัวแข็งเมื่อตาย |
สภาวะที่กล้ามเนื้อแข็งเกร็ง
(physiologic contracture) เกิดขึ้นเนื่องจากการมี ATP อยู่ในเส้นใยกล้ามเนื้อในปริมาณต่ำมากอันเนื่องมาจากการออกกำลังกายอย่างหนัก
หรือภาวะที่มีการอดอาหารอย่างรุนแรง และเกิดขึ้นได้เองโดยไม่ต้องมีสัญญาณประสาทมากระตุ้น
การที่กล้ามเนื้อมี ATP ไม่เพียงพอทำให้กระบวนการปั๊มแคลเซียมไอออนกลับเข้าสู่ร่างแหซาร์โคพลาสมิกเกิดขึ้นได้ไม่ดี
ทำให้มีแคลเซียมไอออนสะสมอยู่ในซาร์โคพลาซึมในปริมาณสูง ส่งผลให้ไมโอซินเข้าเกาะกับแอกทิน
เมื่อ ATP ในเส้นใยกล้ามเนื้อหมดไป ทำให้สะพานเชื่อมระหว่างแอกทิน
และไมโอซินไม่สามารถหลุดออกจากกัน ในที่สุดกล้ามเนื้อทั้งชิ้นจะแข็งเกร็งไม่สามารถหดตัวหรือคลายตัวได้
สภาวะเช่นนี้เห็นได้ชัดในคนที่เสียชีวิตไปแล้วหลายๆ ชั่วโมง
และมีคำเรียกเฉพาะสำหรับร่างที่หมดลมไปแล้วว่าไรกอร์มอร์ทิส
(rigor mortis)
|