








| นาโนเทคโนโลยี |
||
.jpg) |
||
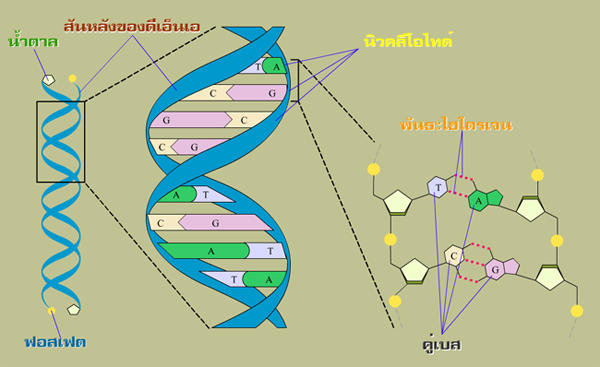
กรดนิวคลีอิค (nucleic acid) เป็นหน่วยประกอบพื้นฐานที่ทำหน้าที่เป็นตัวกำหนดลักษณะของสิ่งมีชีวิต โดยเป็นพอลิเมอร์ของมอนอเมอร์โมเลกุล นิวคลีโอไทด์ ที่เชื่อมต่อเรียงตัวกันเป็นสายอย่างเป็นลำดับ จนได้เป็นสายพอลินิวคลีโอไทด์ (ซึ่งอาจจะมากถึง 100 ล้านนิวคลีโอไทด์ในหนึ่งโมเลกุลกรดนิวคลีอิค)่ โดยที่แต่ละมอนอเมอร์ของนิวคลีโอไทด์เชื่อมต่อกันด้วยพันธะฟอสโฟไดเอสเทอร์ (phospho-diester bond) ซึ่งหมู่ฟอสเฟตที่เป็นส่วนประกอบภายในพันธะจะเชื่อมโยงระหว่างหมู่ไฮดรอกซิลที่ตำแหน่งที่ 5´ ของนิวคลีโอไทด์โมเลกุลที่หนึ่งกับหมู่ไฮดรอกซิลที่ตำแหน่งที่ 3´ ของนิวคลีโอไทด์โมเลกุลถัดไป ทำให้นิวคลีโอไทด์จึงมีโครงสร้างสันหลังของแต่ละสายเกลียวเป็นฟอสเฟตและน้ำตาล โดยมีแขนงข้างภายในสายเกลียวเป็นเบส |
||
|
||
กรดนิวคลีอิค 2 ชนิดที่เป็นที่รู้จักกันดีโดยทั่วไป ได้แก่ กรดไรโบนิวคลีอิคหรืออาร์เอ็นเอ (ribonucleic acid : RNA) และกรดดีออกซีไรโบนิวคลีอิคหรือดีเอ็นเอ (deoxyribonucleic acid : DNA) ซึ่งทั้งสองมีบทบาทสำคัญในกระบวนการสร้างระดับนาโนที่เกิดขึ้นในโครงสร้างของโปรตีน |
||
ดีเอ็นเอเป็นโมเลกุลพอลิเมอร์ทางชีวภาพที่มีลักษณะแตกต่างกัน 4 ชนิด (อาร์เอ็นเอก็เช่นเดียวกัน) โดยส่วนใหญ่ที่พบในธรรมชาติจะมีลักษณะเป็น โครงสร้างเกลียวคู่ ซึ่งเป็นโครงสร้างที่มีความเสถียรที่สุด โดยเกลียวคู่ของดีเอ็นเอที่พบมากที่สุดในธรรมชาติจะเป็นโครงสร้างระดับนาโน ซึ่งมีเส้นผ่านศูนย์กลางประมาณ 2 นาโนเมตร เบสที่อยู่เป็นแขนงข้างแต่ละตัวอยู่ห่างกัน 0.34 นาโนเมตร การเวียนหนึ่งรอบของเกลียวคู่ (ที่มีลำดับเบสอยู่ 10 คู่) คิดเป็นระยะทางได้ 3.4 นาโนเมตร |
||
|
||
โครงสร้างของกรดนิวคลีอิกถูกกำหนดโดยการจับคู่ของเบสในแต่ละนิวคลีโอไทด์ เบสที่อยู่ในแต่ละนิวคลีโอไทด์มีความสามารถที่จะเลือกจับคู่อย่างเฉพาะเจาะจงกับเบสอีกตัวหนึ่ง และเชื่อมต่อกันด้วยพันธะไฮโดรเจน นำไปสู่การเป็นโครงสร้างที่มีลักษณะเป็นเกลียวคู่ของดีเอ็นเอ ซึ่งโครงสร้างเกลียวคู่ของดีเอ็นเอที่พบได้ในธรรมชาติและเป็นที่รู้จักกันดีมี 3 ชนิด ได้แก่ เอ-ดีเอ็นเอ (A-DNA), บี-ดีเอ็นเอ (B-DNA) และ ซี-ดีเอ็นเอ (Z-DNA) โดยที่แต่ละชนิดมีความแตกต่างกันทั้งระยะทางในหนึ่งรอบเกลียว จำนวนเบสต่อรอบเกลียว มุมของเบสที่ทำกับแกนของเกลียว และขนาดของโครงสร้าง (ภาพนิ่งด้านบนเป็นโครงสร้างของบี-ดีเอ็นเอที่มีขนาดเส้นผ่านศูนย์กลางประมาณ 2 นาโนเมตร และมีเบสที่เป็นแขนงข้างแต่ละตัวอยู่ห่างกัน 0.34 นาโนเมตร ส่วนเอ-ดีเอ็นเอและซี-ดีเอ็นเอมีขนาดเส้นผ่านศูนย์กลางอยู่ในช่วงระหว่าง 1.8 - 2.3 นาโนเมตร และมีเบสที่เป็นแขนงข้างแต่ละตัวอยู่ห่างกันในช่วงระหว่าง 0.321 - 0.337 นาโนเมตร) สายพอลินิวคลีโอไทด์นี้มีคุณสมบัติของความยืดหยุ่นมากกว่าสายโพลีเปปไทด์ ดังนั้นแล้วกรดนิวคลีอิคจึงสามารถที่จะปรับเปลี่ยนโครงรูปได้อย่างหลากหลายด้วยเช่นกัน อย่างเช่นโครงสร้าง ดีเอ็นเอทั้ง 3 ชนิดนี้สามารถที่จะเกิดการเปลี่ยนแปลงโครงรูปได้ ขึ้นอยู่กับสภาพแวดล้อมที่เป็นตัวทำละลายของสารละลาย |
||
|
||
เบสทั้ง 4 ชนิดที่ถูกใช้ในการสร้างโครงสร้างดีเอ็นเอ ได้แก่ เบสอะดีนีน (adenin : A) เบสกัวนีน (guanine : G) เบสไซโทซีน (cytosine : C) และเบสไทมีน (thymine : T) (แต่สำหรับอาร์เอ็นเอจะคล้ายคลึงกัน ต่างกันตรงที่เปลี่ยนจากเบสไทมีนเป็นเบสยูเรซิล) โดยเบสทั้ง 4 ชนิดนี้มีคุณสมบัติทางเคมีที่คล้ายคลึงกันมาก แต่ก็มีความแตกต่างในการกำหนดตัวรับและตัวให้กับพันธะไฮโดรเจนที่อยู่รอบๆ ขอบของแต่ละเบส การเข้าคู่กันของเบสตามวิธีของวัตสัน-คริกเกิดขึ้นใน 2 ลักษณะ ได้แก่ คู่เบสระหว่างเบสอะดีนีนกับเบสไทมีน (A กับ T) และคู่เบสระหว่างเบสไซโทซีนกับเบสกัวนีน (C กับ G) ซึ่งทั้งสองคู่เบสนี้ยินยอมให้เกิดการก่อตัวของโครงสร้างที่เป็นเกลียวคู่ (การจับคู่ของเบสแบบอื่นๆ ก็เป็นไปได้ แต่จะไม่ทำให้เกิดเป็นดีเอ็นเอลักษณะเกลียวคู่เช่นนี้) การเรียงลำดับของคู่เบสจำนวนมหาศาลสลับไปมาบนสายเกลียวพอลินิวคลีโอไทด์นั้น เป็นเสมือนรหัสคำสั่งของกระบวนการทั้งมวลที่เกิดขึ้นในโลกชีวภาพ ซึ่งมันคือรหัส นาโนของข้อมูลทางพันธุกรรมที่จะถูกทำสำเนา ถูกอ่าน และถูกส่งผ่านอย่างเฉพาะเจาะจงจากสายเกลียวหนึ่งไปสู่อีกสายเกลียวหนึ่ง จากการทำงานของเอนไซม์ หลายชนิดด้วยกันซึ่งเกิดขึ้นในกระบวนการของการจำลองดีเอ็นเอ (DNA replication) จากนั้นก็รหัสข้อมูลระดับนาโนของดีเอ็นเอก็จะถูกส่งผ่านไปสู่ อาร์เอ็นเอ และไปสู่ไรโบโซมซึ่งเป็นเครื่องประกอบระดับนาโนที่ทำหน้าที่ในการสังเคราะห์โปรตีน เพื่อนำไปสร้างโครงสร้างชีวภาพระดับนาโนอันเป็นรากฐานของสิ่งมีชีวิตทั้งหลายอย่างมากมายต่อไป อย่างเช่น โปรตีน เอนไซม์ และฮอร์โมนต่างๆ |
||
|
||
จากโครงสร้างของกรดนิวคลีอิกจะเห็นได้ว่า กรดนิวคลีอิกมีแนวทางในการสร้างโครงสร้างในระดับอะตอมหรือโมเลกุล (ซึ่งอยู่ในช่วงอาณาจักรระดับนาโน) ที่คล้ายกันกับโปรตีน โดยเริ่มจากการที่มอนอเมอร์โมเลกุลนิวคลีโอไทด์แต่ละตัวถูกเชื่อมต่อกันโดยอาศัยพันธะฟอสโฟไดเอสเทอร์ ทำให้เกิดเป็นสายพอลิเมอร์ของนิวคลีโอไทด์ จนสุดท้ายได้เป็นโครงสร้างสายพอลินิวคลีโอไทด์ อย่างเช่น ถ้าเป็นดีเอ็นเอก็จะเกิดเป็นสายพอลิดีออกซีไรโบนิวคลีอิก หรือถ้าเป็นอาร์เอ็นเอก็จะได้สายพอลิไรโบนิวคลีอิกเช่นเดียวกัน และโดยการเชื่อมต่อพันธะฟอสโฟไดเอสเทอร์ของแต่ละน้ำตาลในนิวคลีโอไทด์ และความสามารถที่จะจัดตัวเองให้เบสของทั้งสองสายที่จับคู่กันต้องวางเกือบอยู่ในระนาบเดียวกัน เพื่อให้เบสแต่ละคู่สามารถเชื่อมต่อกันด้วยพันธะไฮโดรเจนได้ ทำให้กรดนิวคลีอิกสามารถจัดตัวเองเข้าเป็นโครงสร้างแบบเกลียวคู่ที่มีความเสถียร และมีความหลากหลายได้ อีกทั้งกรดนิวคลีอิกยังสามารถที่จะประกอบตัวเองได้ โดยการจับคู่ของคู่เบสที่ถูกต้องภายในดีเอ็นเอ ซึ่งก็คือระหว่างเบสอะดีนีน (A) กับเบสไทมีน (T) และเบสกัวนีน (G) กับเบสไซโทซีน (C) เพื่อใช้ในการจัดเก็บรหัสข้อมูลระดับนาโนของพันธุกรรม เพื่อนำไปสร้างโครงสร้างระดับนาโนต่างๆ ต่อไป (หรือระหว่างเบสไทมีน (A) กับเบสยูเรซิล (U) และเบสกัวนีน (G) กับเบสไซโตซีน (C) ภายในอาร์เอ็นเอเช่นเดียวกัน) |
||
-edit.jpg) |
||
.jpg) |
||
ไรโบโซม (ribosome) เป็นอวัยวะย่อยขนาดเล็กภายในเซลล์ที่ไม่มีเยื่อหุ้ม มีรูปร่างลักษณะเป็นก้อน ประกอบไปด้วยโปรตีนและอาร์เอ็นเอที่มีสัดส่วนเท่ากันโดยน้ำหนัก ซึ่งไรโบโซมที่เกาะอยู่ที่ผิวนอกของเอนโดพลาสมิค เรติคิวลัมผิวขรุขระ (Rough Endoplasmic Reticulum: RER) จะทำหน้าที่เป็นแหล่งสร้างโปรตีนสำหรับไว้ใช้เป็นองค์ประกอบของเยื่อหุ้มเซลล์และส่งออกไปใช้ภายนอกเซลล์ และยังมีไรโบโซมอิสระที่ไม่เกาะอยู่กับเอนโดพลาสมิค เรติคิวลัม (Endoplasmic Reticulum: ER) โดยจะกระจายอยู่ในไซโทซอล (Cytosol) ทำหน้าที่สร้างโปรตีนสำหรับไว้ใช้ภายในเซลล์และยังทำหน้าที่สร้างฮีโมโกลบิน ซึ่งจะพบมากในเซลล์เม็ดเลือดแดงที่มีช่วงอายุน้อย |
||
|
||
ไรโบโซม เป็นเสมือนเครื่องประกอบระดับนาโน (nanoassembler) ที่มีขนาดเส้นผ่านศูนย์กลางเพียง 11 นาโนเมตรเท่านั้น และทำหน้าที่สังเคราะห์โปรตีนตามข้อมูลของเอ็ม-อาร์เอ็นเอ (mRNA) ที่ได้รับมาจากดีเอ็นเอ (DNA) โดยไรโบโซมจะประกอบด้วยหน่วยย่อย 2 หน่วยย่อย ได้แก่ หน่วยย่อยขนาดใหญ่ (large subunit) และหน่วยยย่อยขนาดเล็ก (small subunit) โดยบนโครงสร้างนาโนของไรโบโซมจะมีตำแหน่งที่โมเลกุลของที-อาร์เอ็นเอ (tRNA) สามารถจับตัวอยู่ได้ 2 ตำแหน่ง คือ P-site (peptidyl site) และ A-site (amino acyl site) ซึ่งตำแหน่ง P-site เป็นที่จับตัวของ tRNA ที่มีสายเปปไทด์เกาะอยู่ ส่วน A-site เป็นตำแหน่งที่จับตัวของ tRNA ที่มีกรดอะมิโนเกาะอยู่ ลำดับของกรดอะมิโนของโปรตีนต้องอาศัยการจับคู่อย่างจำเพาะระหว่างโคดอนของโมเลกุล mRNA กับแอนติโคดอนของ tRNA ซึ่ง tRNA นั้นจะเป็นตัวนำพากรดอะมิโนที่ถูกต้องเข้ามาสู่โครงสร้างของไรโบโซม |
||
|
||
กระบวนการสังเคราะห์โปรตีนเป็นกระบวนการที่มีความซับซ้อน เพราะรหัสทางพันธุกรรมเป็นรหัสข้อมูล (informational code) ซึ่งไม่ได้เป็นรหัสทางเคมี (chemical code) แต่อย่างใด โดยที่ การคัดลอกรหัสข้อมูล (transcription) จากดีเอ็นเอต้นแบบไปยังอาร์เอ็นเอใช้รหัสทางเคมีในการดำเนินการ โดยเกิดขึ้นเมื่อลำดับของเบสแต่ละตัวภายในดีเอ็นเอจับตัวเข้าคู่กันกับเบสของอาร์เอ็นเอ โดยมีเอนไซม์อาร์เอ็นเอ โพลิเมอเรส (RNA polymerase) เป็นตัวช่วยในการเชื่อมต่อกันระหว่างเบสของอาร์เอ็นเอ ให้เข้ากับสายเกลียวของดีเอ็นเอได้อย่างเหมาะสม ส่วนในขั้นตอนของการแปลรหัสข้อมูล (translation) ของอาร์เอ็นเอไปเป็นโปรตีนนั้นไม่ได้ใช้ปฏิสัมพันธ์ทางเคมีระหว่างอาร์เอ็นเอกับกรดอะมิโน แต่ทว่าเป็นการแปลความหมายของภาษาทางเคมีให้ไปเป็นอีกภาษาหนึ่งที่มีความหมายเหมือนกัน (ซึ่งก็คือเป็นรหัสข้อมูลนั่นเอง) โดยที่มี tRNA เป็นโมเลกุลที่คอยเป็นผู้ทำหน้าที่ในการแปลความหมายข้อมูล (สามารถเรียกได้ว่า tRNA เป็นพจนานุกรมสำหรับการแปลรหัสข้อมูลนี้) |
||
ไรโบโซมทำหน้าที่เปรียบเสมือนสถานที่หรือโรงงานสำหรับประกอบโปรตีนโดยเฉพาะ เป็นจักรกล นาโนที่ใช้พลังงานทางเคมี คือ GTP ในการขับเคลื่อนเพื่อดำเนินการสังเคราะห์โปรตีน โดยในกระบวนการสังเคราะห์โปรตีน เริ่มต้นจากไรโบโซมหน่วยย่อยขนาดเล็กจะเข้าจับตัวกับโมเลกุลของ mRNA โคดอน (codon) เริ่มต้นจะถูกแปลรหัสเป็นกรดอะมิโนคือ AUG (โคดอนเป็นกลุ่มของเบส 3 ตัวที่เรียงตัวกันตามลำดับบนสาย mRNA โดย 1 โคดอนของ 3 นิว คลีโอไทด์จะถูกใช้สำหรับกรดอะมิโน 1 ตัว) จากนั้นโมเลกุลของ tRNA สำหรับโคดอนแรกจะนำกรดอะมิโนตัวแรกคือ Met (ซึ่งมีแอนติโคดอนเป็น UAC) เข้ามาจับกับโคดอน AUG ของโมเลกุล mRNA ที่ตำแหน่ง P-site และในขณะเดียวกันหน่วยย่อยขนาดใหญ่ของไรโบโซมก็จะเข้าไปรวมตัวกับหน่วยย่อยขนาดเล็ก จนเกิดเป็นไรโบโซมผู้ประกอบระดับนาโนที่สมบูรณ์ที่พร้อมจะดำเนินการสังเคราะห์โปรตีน [ซึ่งการจับตัวกันระหว่างไรโบโซม mRNA และ tRNA เกิดเป็นคอมเพล็กซ์เริ่มต้น (initial complex) สำหรับการสร้างโปรตีนนี้ จำเป็นต้องใช้โปรตีนแฟคเตอร์อีกชนิดหนึ่ง คือ IF หรือ initial factor] ต่อจากนั้นในตำแหน่ง A-site ที่ว่างอยู่ของไรโบโซมนั้นก็จะมีโมเลกุลของ tRNA ที่นำกรดอะมิโนตัวต่อไปเข้ามาจับ ซึ่ง amino acyl-tRNA ตัวใหม่ที่เข้ามาจับนี้จะต้องมีแอนติโคดอนบนโมเลกุล tRNA ที่จำเพาะเจาะจงกับโคดอนบนโมเลกุล mRNA ทำให้เกิดการสร้างคู่เบสได้ ปฏิกิริยาต่อไปก็คือการสร้างพันธะเปปไทด์ระหว่างหมู่แอลฟาอะมิโนของ amino acyl-tRNA ตัวใหม่ที่บริเวณ A-site และหมู่คาร์บอกซิลของ peptidyl-tRNA ที่บริเวณ P-site ซึ่งจะทำให้เกิดการเชื่อมต่อเปปไทด์ที่ยาวขึ้นกับ peptidyl-tRNA ของ A-site จากนั้นโมเลกุล peptidyl-tRNA ที่บริเวณ P-site ก็จะเป็น tRNA อิสระ (คือไม่มีกรดอะมิโนเกาะอยู่) หลังจากนั้นไรโบโซมก็จะเคลื่อนตัวไปทางด้านปลาย 3´ ของโมเลกุล mRNA เป็นระยะ 1 โคดอน มีผลทำให้ peptidyl-tRNA ที่อยู่บริเวณ A-site เปลี่ยนไปอยู่ที่บริเวณ P-site และโมเลกุล tRNA อิสระหลุดออกจากไรโบโซม ส่วนตำแหน่ง A-site ก็จะว่างอีกครั้งเพื่อรอรับโมเลกุล amino acyl-tRNA ตัวใหม่เข้ามา แล้วสร้างพันธะเปปไทด์ระหว่างหมู่คาร์บอกซิลที่ปลาย C ของสายเปปไทด์กับหมู่อะมิโนของกรดอะมิโนตัวใหม่ที่เข้ามาอีกรอบ แล้วก็จะเกิดวงจรการสังเคราะห์สายโพลีเปปไทด์อย่างนี้ไปเรื่อยๆ จากปลายด้าน N ไปสู่ปลายด้าน C จนกว่าไรโบโซมจะเคลื่อนตัวไปถึงรหัสยุติ (stop codon) บนสายโมเลกุล mRNA ซึงเมื่อมาถึงจุดนี้ก็จะมีการสลายพันธะระหว่างสายโพลิเปปไทด์ที่กำลังถูกสร้างขึ้นกับโมเลกุล tRNA ที่บริเวณ P-site ทำให้สายโพลีเปปไทด์หลุดออกมาจากไรโบโซม แล้วไรโบโซมทั้งสองหน่วยย่อยก็จะแยกกออกจากัน (ซึ่งในขั้นตอนสุดท้ายนี้ต้องอาศัยโปรตีนแฟคเตอร์อีกตัวหนึ่ง คือ RS หรือ release factor) สุดท้ายก็จะได้สายโพลีเปปไทด์หรือโปรตีนนั่นเอง |
||
.jpg)

.jpg)
.jpg)







 - มอเตอร์เชิงเส้นของไมโครทิวบิล(ไคเนซิน-ไมโครทิวบิล)]")
]")
 - ช่องเข้าออกเยื่อหุ้มเซลล์ควบคุมด้วยความต่างศักย์ไฟฟ้า(Voltage-gated ion channel)]")



