








| นาโนเทคโนโลยี |
|
.jpg) |
|
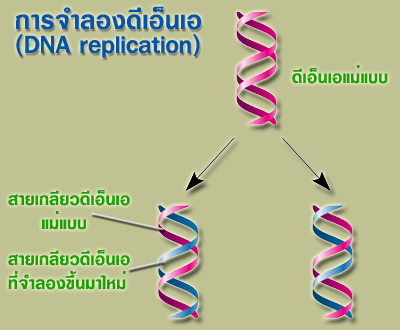
การจำลองดีเอ็นเอเป็นกระบวนการทางธรรมชาติอันซับซ้อน ก็เพราะว่าดีเอ็นเอเป็นโมเลกุลชีวภาพที่เป็นสารพันธุกรรม และบรรจุข้อมูลทางพันธุกรรมของสิ่งมีชีวิต ดังนั้นกระบวนการของการจำลองดีเอ็นเอ จึงเป็นกระบวนการของการจำลองข้อมูลทางพันธุกรรมด้วยเช่นเดียวกัน ที่เป็นเหมือนกับการถ่ายแบบดีเอ็นเอเดิมทุกประการ เพื่อเป็นการรักษาข้อมูลทางพันธุกรรมที่จะถูกถ่ายทอดไปยังลูกหลานต่อไป |
|
| กระบวนการของการจำลองดีเอ็นเอเป็นแบบกึ่งอนุรักษ์ (semiconservative) โดยสายเกลียวทั้งสองของดีเอ็นเอจะถูกแยกออกจากกันเพื่อทำหน้าที่เป็นแม่แบบ (template) ในการสร้างสายดีเอ็นเอสายใหม่ และโดยการจับคู่เบสตามแบบวัตสัน-คริก จะทำให้ได้ดีเอ็นเอคู่ใหม่ 2 คู่ที่มีลำดับนิวคลีโอไทด์เหมือนดีเอ็นเอแม่แบบทุกประการ |
|
|
|
โดยทั่วไปการจำลองดีเอ็นเอจะเกิดขึ้นเมื่อมีการแบ่งเซลล์ กลไกของการจำลองดีเอ็นเอค่อนข้างจะมีการทำงานที่ซับซ้อน โดยมีเอนไซม์และโปรตีนหลายชนิดมาเกี่ยวข้อง ดังนั้นจึงต้องการการทำงานที่เป็นหน่วยรวม ซึ่งการจำลองดีเอ็นเอจะเริ่มขึ้นที่จุดเริ่มต้นจำเพาะที่เรียกว่าเป็นง่ามการจำลอง (replication folk) โดยที่จะมีเอนไซม์ดีเอ็นเอ เฮลิเคส (DNA helicase) ทำหน้าที่คลายสายเกลียวคู่ของดีเอ็นเอแม่แบบให้แยกสายออกจากกัน โดยการทำลายพันธะไฮโดรเจนระหว่างคู่เบสของดีเอ็นเอ หลังจากนั้นจะมีโปรตีนจำนวนมากมาจับกับดีเอ็นเอสายเดี่ยวแต่ละสาย เพื่อป้องกันไม่ให้สายเดี่ยวของดีเอ็นเอแต่ละสายกลับไปรวมตัวกันเป็นเกลียวคู่ได้อีก ถึงขั้นตอนนี้จะได้ส่วนของดีเอ็นเอที่เป็นสายเดี่ยว 2 สายที่พร้อมที่จะทำหน้าที่เป็นแม่แบบสำหรับการสร้างดีเอ็นเอสายใหม่ |
|
เมื่อดีเอ็นเอสายเกลียวคู่แยกออกจากกันโดยเอนไซม์ดีเอ็นเอ เฮลิเคสแล้ว ส่วนที่อยู่เหนือจุดแยกของสายเกลียวคู่นั้นย่อมจะขดและม้วนตัว เกิดเป็นเกลียวซ้อนเกลียวขึ้น (super coiling) ทำให้เอนไซม์ดีเอ็นเอ เฮลิเคสไม่สามารถที่จะแยกสายเกลียวคู่ของดีเอ็นเอแม่แบบต่อไปได้ ดังนั้นจึงจำเป็นต้องมีเอนไซม์อีกตัวหนึ่งก็คือ เอนไซม์โทโปไอโซเมอเรส (topoisomerase) จะเข้ามาทำหน้าที่คลายเกลียวบริเวณที่เป็นเกลียวซ้อนเกลียวของดีเอ็นเอ โดยการตัดสายของดีเอ็นเอ เพื่อป้องกันการพันกันอย่างยุ่งเหยิง และการขมวดเป็นปมของสายดีเอ็นเอในระหว่างการจำลองตัวดีเอ็นเอขึ้นมาอีก |
|
| ในขณะที่ดีเอ็นเอแม่แบบคลายเกลียวแยกออกจากกัน การสร้างดีเอ็นเอสายใหม่ขึ้นมาจะเกิดขึ้นใน 2 ลักษณะ คือ สายหนึ่งจะถูกสร้างขึ้นในลักษณะที่ต่อเนื่องกันไปเป็นสายยาว และสร้างไปในทิศทางเดียวกับการเคลื่อนที่ของง่ามการจำลอง ซึ่งเรียกว่าสายนำ (leading strand) โดยในสายนำนี้จะถูกสร้างขึ้นโดยการทำงานของเอนไซม์ดีเอ็นเอ โพลิเมอรเรส III (DNA polimerase III) เพียงอย่างเดียว ส่วนอีกลักษณะหนึ่งจะเกิดขึ้นโดยที่อีกสายหนึ่งของดีเอ็นเอที่ถูกแยกแล้ว จะถูกสร้างขึ้นในลักษณะเป็นช่วงสั้นๆ เป็นระยะๆ อย่างไม่ต่อเนื่อง (ซึ่งช่วงสั้นๆ ของเส้นนั้นเรียกว่าโอกาซากิ แฟรกเมนท์ (okazaki fragments)) และมีทิศทางตรงกันข้ามกับการเคลื่อนที่ของง่ามการจำลอง ซึ่งเรียกว่า สายตาม (lagging strand) โดยในสายตามนี้การสร้างดีเอ็นเอสายใหม่จะเริ่มขึ้นด้วยการสร้างอาร์เอ็นเอ ไพรเมอร์ (RNA primer) สั้นๆ ขึ้นก่อนโดยการทำงานของเอนไซม์อาร์เอ็นเอ ไพรเมส (RNA primase) จากนั้นดีเอ็นเอสายใหม่จะถูกสร้างต่อจากอาร์เอ็นเอ ไพรเมอร์อีกทีหนึ่ง การสร้างสายใหม่ในแต่ละช่วงเกิดขึ้นโดยการทำงานของเอนไซม์ดีเอ็นเอ โพลิเมอรเรส III ในทิศทาง 5´ ไปหา 3´ เสมอ ซึ่งเป็นทิศทางที่สวนทางกับสายดีเอ็นเอแม่แบบ ต่อมาเอนไซม์ดีเอ็นเอ โพลิเมอเรส I (DNA polimerase I) จะทำหน้าที่ตัดอาร์เอ็นเอ ไพรเมอร์ออกไป และสร้างสายของดีเอ็นเอเข้าไปแทนที่ สุดท้ายดีเอ็นเอเส้นสั้นๆ ก็จะถูกเชื่อมต่อกันให้เป็นดีเอ็นเอสายยาวได้โดยการทำงานของเอนไซม์ดีเอ็นเอ ไลเกส (DNA ligase) |
|
| เมื่อสิ้นสุดกระบวนการจำลองดีเอ็นเอแล้ว ก็จะได้ดีเอ็นเอเกลียวคู่ 2 คู่ โดยแต่ละสายจะประกอบไปด้วยแม่บแบบ 1 สาย และสายที่สร้างขึ้นใหม่อีก 1 สาย โดยที่ดีเอ็นเอเกลียวคู่ที่เกิดใหม่นั้นจะมีลำดับนิวคลีโอไทด์เหมือนกับดีเอ็นเอแม่แบบทุกประการ |
|
.jpg) |
|
| เซลล์เป็นหน่วยพื้นฐานของสิ่งมีชีวิตทุกชนิด และเซลล์ของสิ่งมีชีวิตจะมีเยื้อหุ้มล้อมรอบเรียกว่า เยื่อหุ้มพลาสมา (plasma membrane) โดยเป็นเยื่อหุ้มที่มีลักษณะเป็นไขมัน 2 ชั้น ที่หันส่วนที่ชอบน้ำออกด้านนอกและส่วนที่ไม่ชอบน้ำเข้าด้านใน (หรือไบเลเยอร์นั่นเอง) จึงทำให้มีผลต่อการควบคุมการผ่านเข้าออกของสารที่จะผ่านชั้นไขมันนี้ โดยสารจำพวกไฮโดรโฟบิก เช่น |
|
 |
|
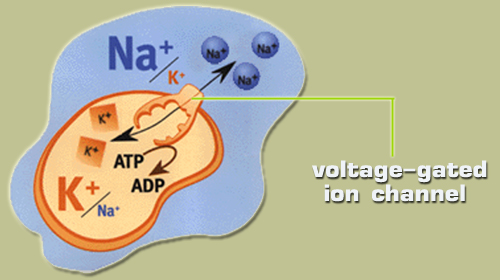
ช่องเข้าออกเยื่อหุ้มเซลล์ควบคุมด้วยความต่างศักย์ไฟฟ้าเป็นช่องที่ฝังอยู่กับเยื่อหุ้มเซลล์ สำหรับควบคุมการผ่านเข้าออกภายในเซลล์ของไอออน โดยที่การทำงานของโครงสร้างนาโนของช่องเข้าออกเยื่อหุ้มเซลล์ควบคุมด้วยความต่างศักย์ไฟฟ้านี้ จะเกิดการทำงานขึ้นก็ต่อเมื่อถูกกระตุ้นโดยศักย์ไฟฟ้าที่แตกต่างกัน ระหว่างภายในเซลล์และภายนอกเซลล์ที่อยู่โดยรอบช่องนี้นั่นเอง โดยเมื่อเซลล์อยู่ในสภาะสมดุล ซึ่งก็คือสภาวะที่เซลล์มีความเข้มข้นของโพแทสเซียมไอออนภายในเซลล์มากกว่าภายนอกเซลล์ ( |
|
กระบวนการแลกเปลี่ยนโซเดียมไอออนและโพแทสเซียมไอออน (โซเดียมโพแทสเซียมไอออนปั๊ม) เป็นกระบวนการทำงานของโครงสร้างนาโนช่องเข้าออกเยื่อหุ้มเซลล์ควบคุมด้วยความต่างศักย์ไฟฟ้าที่ประกอบด้วยขั้นตอนทั้งหมด 6 ขั้นตอนดังนี้ 1. โปรตีนขนส่งเอทีพีเอสมีตำแหน่งว่างที่เหมาะสมจะจับโซเดียมไอออนจำนวน 3 ไอออนจากทางด้านในเซลล์ 2. โมเลกุลเอทีพีเกิดไฮโดรไลซิส (hydrolysis) แตกตัวเป็นโมเลกุลเอดีพีและฟอสเฟต จากนั้นโมเลกุลหมู่ฟอสเฟตจับตัวที่โปรตีนขนส่ง 3. เกิดการเปลี่ยนแปลงโครงร่างของโปรตีนขนส่ง ทำให้โซเดียมไอออนถูกปล่อยออกนอกเซลล์ 4. โปรตีนขนส่งมีตำแหน่งว่างเหมาะสมที่จะจับโพแทสเซียมไอออน ดังนั้นโพแทสเซียมไอออนที่อยู่ด้านนอกเซลล์จำนวน 2 ไอออนจะเข้าจับที่โปรตีนขนส่ง 5. โปรตีนขนส่งเกิดดีฟอสโฟรีเลชัน (dephosphorylation) ทำให้หมู่ฟอสเฟตถูกปล่อย ส่งผลให้เกิดการเปลี่ยนแปลงโครงร่างของโปรตีนขนส่งอีกครั้ง 6. โปรตีนขนส่งพร้อมที่จะปล่อยโพแทสเซียมไอออนเข้าสู่ภายในเซลล์ ดังนั้นในกระบวนการของการแลกเปลี่ยนไอออนของโครงสร้างนาโนช่องเข้าออกเยื่อหุ้มเซลล์ควบคุมด้วยความต่างศักย์ไฟฟ้านั้น จะเกิดการแลกเปลี่ยนกันระหว่างโซเดียมไอออน 3 ไอออนจากภายในเซลล์ถูกส่งออกไปนอกเซลล์ และโพแทสเซียมไอออน 2 ไอออนจากภายนอกเซลล์ถูกส่งให้เข้ามาภายในเซลล์ |







 - มอเตอร์เชิงเส้นของไมโครทิวบิล(ไคเนซิน-ไมโครทิวบิล)]")
]")
_38.jpg "singleclickstop=[on] header=[นาโนเทคโนโลยีชีวภาพในระบบธรรมชาติ]
body=[หน่วยรวมที่ช่วยในการจำลองดีเอ็นเอ(DNA replication complex) - ช่องเข้าออกเยื่อหุ้มเซลล์ควบคุมด้วยความต่างศักย์ไฟฟ้า(Voltage-gated ion channel)]")



