








นาโนเทคโนโลยี |
||||||
.jpg) |
||||||
| เซลล์เป็นสิ่งมหัศจรรย์ของสิ่งมีชีวิตทุกชนิด ซึ่งภายในเซลล์ก็จะมีโปรตีนต่างๆ ที่เป็นจักรกลนาโนทำหน้าที่เกี่ยวกับกลไกต่างๆ ภายในเซลล์ แต่บางครั้งเซลล์ก็อาจจะพบกับปัญหาในการสร้างจักรกลนาโนโปรตีนเหล่านั้นได้ และเป็นที่ทราบกันแล้วว่าโปรตีนเป็นสายโพลีเปปไทด์ที่ถูกสังเคราะห์ขึ้นโดยผู้ประกอบระดับนาโนที่เป็นโมเลกุล นั่นก็คือไรโบโซม โดยกระบวนการในการสังเคราะห์นั้นเกิดขึ้นจากการทำงานร่วมกันระหว่างโมเลกุล tRNA ที่ทำหน้าที่เป็นตัวแปลรหัสจากลำดับเบสที่อยู่บนสาย mRNA มาเป็นกรดอะมิโนแต่ละชนิด โดย tRNA นี้จะเป็นตัวนำกรดอะมิโนที่ถูกต้องเข้ามาเพื่อเชื่อมต่อกันเป็นสายโพลีเปปไทด์ตรงไรโบโซม โดยในกระบวนการสังเคราะห์นั้นจะมีไรโบโซมหลายๆ ตัวทำงานพร้อมกันบนสาย mRNA สายเดียวกัน จนกระทั่งสุดท้ายได้เป็นสายโพลีเปปไทด์ออกมา จากการที่มีไรโบโซมหลายๆ ตัวทำงานพร้อมๆ กันนี้ ทำให้สายโพลีเปปไทด์หลายสายถูกสร้างขึ้นมา ผลที่อาจจะเกิดขึ้นก็คือสายโพลีเปปไทด์เหล่านี้อาจเกิดการรวมตัวเข้าด้วยกัน หรือสายโพลีเปปไทด์ที่เพิ่งถูกสร้างขึ้นอาจเกิดการม้วนตัวผิดรูปแบบไป ซึ่งโครงสร้างที่ผิดไปนี้ทำให้เป็นโปรตีนที่ไม่สามารถทำงานได้ (โดยปกติสายโพลีเปปไทด์แต่ละสายจะม้วนตัวเข้าเป็นโครงสร้างแบบต่างๆ) |
||||||
|
|
||||||
แต่จากการศึกษาทางวิทยาศาสตร์ที่ผ่านมาพบว่า ในธรรมชาตินั้นปัญหาที่เกิดขึ้นนี้ถูกจัดการได้โดยการทำงานของโปรตีนชนิดหนึ่งที่เรียกว่า แชพเพอโรน (chaperone) ซึ่งมีอยู่มากกว่า 50 ชนิด แต่ชนิดหนึ่งที่มีการศึกษาเกี่ยวกับโครงสร้างและบทบาทอย่างจริงจังนั้น คือ แชพเพอโรนิน (chaperonin) โดยพบว่า เป็นโปรตีนที่ทำหน้าที่ป้องกันการม้วนพับของสายโพลีเปปไทด์ที่เพิ่งถูกสร้างไม่ให้ม้วนผิดรูปแบบ ป้องกันการรวมตัวกันของสายโพลีเปปไทด์แล้วเกิดเป็นโปรตีนที่ไม่สามารถทำงานได้ และช่วยให้สาย โพลีเปปไทด์ม้วนพับได้เป็นโครงสร้างที่ถูกต้อง |
||||||
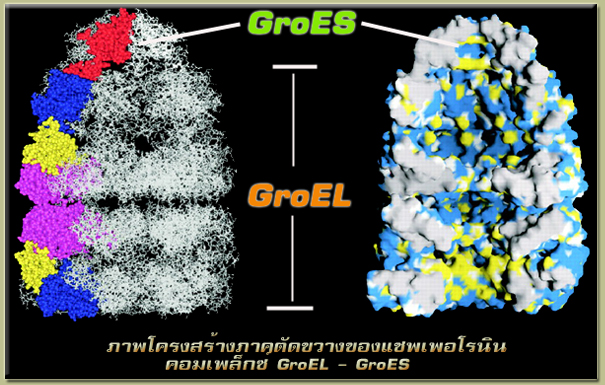
| โปรตีนแชพเพอโรนินเป็นหน่วยรวมของ GroEL กับ GroES (GroEL/GroES complex) พบได้ในแบคทีเรีย E. Coli โดยจากการศึกษาพบว่าหน่วยรวมของ GroEL กับ GroES มีส่วนหนึ่งของโครงสร้างที่มีลักษณะเป็นกรง (cage) ที่มีขนาดระดับนาโน (โดยมีขนาดกว้าง 8 นาโนเมตรและยาว 8.5 นาโนเมตรเท่านั้น ดังนั้นจึงเรียกได้ว่าเป็นกรงนาโน (nanocage) ที่ทำหน้าที่ช่วยเหลือในการม้วนตัวของสายโพลีเปปไทด์ที่เกิดการม้วนตัวผิดรูปแบบไป (อันจะส่งผลให้เกิดเป็นโปรตีนที่ไม่สามารถทำงานได้) ให้สามารถม้วนตัวได้อย่างถูกต้อง โดยเมื่อสายโพลีเปปไทด์เคลื่อนที่เข้าไปภายในโครงสร้างกรงนาโนของแชพเพอโรนินแล้ว จะทำให้สายโพลีเปปไทด์ที่ม้วนพับผิดรูปนั้นคลายแยกออกจากกัน และสามารถม้วนตัวต่อไปอย่างถูกต้องได้ภายในกรงนาโนนี้ นอกจากนั้นยังพบว่าสายโพลีเปปไทด์ที่เข้ามาภายในกรงนาโนนี้สามารถที่จะม้วนตัวได้เร็วมากขึ้นถึง 3-4 เท่าเลยทีเดียว |
||||||
| โครงสร้างโมเลกุล GroEL – GroES นั้นประกอบไปด้วยโมเลกุล GroEL, โมเลกุล GroES และเอนไซม์เอทีพีเอส (ATPase) ซึ่งโมเลกุล GroEL ประกอบด้วยวงแหวน 2 วง ซึ่งแต่ละวงแหวนประกอบขึ้นมาจากหน่วยย่อยทั้งสิ้น 7 หน่วยย่อยที่จัดเรียงตัวกัน และโมเลกุล GroES เป็นวงแหวนเดี่ยวรูปโดมที่ประกอบขึ้นมาจากหน่วยย่อย 7 หน่วยเช่นเดียวกัน โดยที่วงแหวนแต่ละวงของ GroEL จะสามารถต่อตัวเข้าด้วยกันได้นั้นต้องอาศัยให้วงแหวนวงใดวงหนึ่งถูกจับตัวอยู่กับโมเลกุลของเอทีพี (ATP) หรือเอดีพี (ADP) เสียก่อน หลังจากนั้นโมเลกุล GroES ก็จะเคลื่อนที่เข้าจับกับวงแหวนเดียวกันนั้นของโมเลกุล GroEL ก็จะทำให้เกิดเป็นโครงสร้างของกรงนาโนขึ้นมาได้ |
||||||
 |
||||||
| กลไกการทำงานของหน่วยรวม GroEL – GroES จะเริ่มขึ้นเมื่อสายโพลีเปปไทด์ที่สังเคราะห์เสร็จถูกปล่อยออกมาจากโมเลกุลของไรโบโซม ซึ่งในระหว่างนั้นก็เกิดการม้วนตัวไปแล้วบางส่วน โดยสายโพลีเปปไทด์นั้นก็จะเข้าจับตัวกับด้านหนึ่งที่เป็นปลายเปิดของ GroEL (ในขณะที่อีกด้านหนึ่งถูกปิดคลุมโดยการจับตัวของ GroES ) แต่สายโพลีเปปไทด์จะยังไม่สามารถผ่านเข้ามาสู่ช่องภายในของ GroEL ได้ อันเนื่องจากคุณสมบัติของการไม่ชอบน้ำบริเวณในส่วนที่เป็นปลายเปิดของ GroEL นั่นเอง ต่อจากนั้นเมื่อมีโมเลกุลเอทีพี (ATP ) มาจับตัวกับวงแหวนของ GroEL (ส่วนที่มีสายโพลีเปปไทด์ติดอยู่) ทำให้เกิดการหลุดออกไปของโมเลกุลเอดีพี (ADP) ที่ติดอยู่กับอีกวงแหวนหนึ่ง พร้อมกันนั้นก็ทำให้ GroEL เกิดการเปลี่ยนแปลงโครงรูปไป และทำให้สายโพลีเปปไทด์ที่ติดอยู่ถูกปล่อยให้เลื่อนลงผ่านเข้ามาสู่บริเวณภายในของ GroEL ได้ แล้วสุดท้ายก็จะมี GroES (เป็นโมเลกุลที่มีลักษณะเหมือนเป็นฝาปิดที่เคลื่อนที่ได้) เคลื่อนที่เข้ามาปิดคลุมส่วนที่เป็นปลายเปิดของ GroEL นั้น และจับตัวเข้าด้วยกันอันส่งผลให้เกิดเป็นช่องว่างภายในของ GroEL ที่มีลักษณะเป็นกรง (หรือที่เรียกว่ากรงนาโนนั่นเอง) เพื่อกักเก็บสายโพลีเปปไทด์ไว้ภายใน หลังจากนั้นโมเลกุลเอทีพีที่จับตัวอยู่กับ GroEL ก็จะเกิดการไฮโดรไลซ์ขึ้นกลายเป็นโมเลกุลเอดีพี ในขณะเดียวกันนั้นสายโพลีเปปไทด์ก็จะทำการม้วนตัวอยู่ภายในกรงนาโนของคอมเพล็ก GroEL – GroES นี้เป็นระยะเวลาประมาณ 10 -15 วินาที (จนกระทั่งกรดอะมิโนที่มีหมู่ที่ไม่ชอบน้ำ (hydrophobic) ของสายโพลีเปปไทด์ถูกจัดตัวเข้าเป็นโครงสร้างของโปรตีนเฉพาะอย่างถูกต้องได้) ต่อจากนั้นเมื่อมีโมเลกุลเอทีพีใหม่เข้ามาจับตัวกับวงแหวนอีกวงหนึ่งของ GroEL (คือวงแหวนที่มีด้านหนึ่งเป็นส่วนปลายเปิดอยู่) ก็จะทำให้โมเลกุลเอดีพีที่ติดอยู่กับอีกวงแหวนหนึ่งหลุดออกไป พร้อมทั้งกระตุ้นให้ GroES และสายโพลีเปปไทด์ที่ม้วนตัวอย่างถูกต้องแล้วภายในกรงนาโนก็จะถูกปล่อยออกไป |
||||||
.jpg)
|
||||||
มอเตอร์ที่ใช้ขับเคลื่อนแฟลเจลลัมของแบคทีเรีย (bacterial flagellar motor) เป็นจักรกลนาโนชีวภาพที่สมบูรณ์แบบของธรรมชาติอีกตัวอย่างหนึ่ง ที่ทำหน้าที่ช่วยในการว่ายเพื่อเคลื่อนที่ของแบคทีเรีย โดยการเคลื่อนที่ได้นี้เกิดจากการทำงานของมอเตอร์โดยการหมุนสายยาวที่อยู่ในแฟลเจลลัม (filament) หลายๆ สาย ซึ่งในปัจจุบันเหล่านักวิทยาศาสตร์กำลังทำการศึกษาค้นคว้า เพื่อให้เข้าใจอย่างถ่องแท้เกี่ยวกับการทำงานของมอเตอร์ของแบคทีเรียนี้ และในอนาคตอาจจะมีการนำมาประยุกต์ใช้ทางด้านนาโนเทคโนโลยีในการสร้างมอเตอร์ที่มีขนาดเล็กที่สุด เพื่อนำมาใช้ประโยชน์ต่อไปในวันข้างหน้า
|
||||||
|
||||||
| สำหรับเซลล์โปรคาริโอต ระบบธรรมชาติได้สร้างอวัยวะนอกเซลล์ที่เรียกว่า แฟลเจลลัม (flagellum) ให้กับเซลล์ เช่น แบคทีเรียหรือสเปิร์ม โดยเป็นอวัยวะที่มีมอเตอร์แบบหมุน (rotary motor) ที่มีขนาดเส้นผ่านศูนย์กลางประมาณ 30 นาโนเมตร เพื่อให้แบคทีเรียสามารถที่จะเคลื่อนที่ไปได้ อย่างเช่น แบคทีเรียแซลโมเนลลา (salmonella) และแบคทีเรียเอสเชอริเชีย โคไล (escherichia coli หรือ E. coli) ซึ่งมีขนาดลำตัวประมาณ 1 – 2 ไมครอน ความสามารถในการเคลื่อนที่ของแบคทีเรียนี้ ถูกขับเคลื่อนโดยการหมุนอย่างรวดเร็วของใบพัดแบบเกลียวที่ติดอยู่กับมอเตอร์ขนาดเล็ก ซึ่งเป็นมอเตอร์ที่อยู่ภายในส่วนที่เป็นฐาน (basal body) ที่อยู่บริเวณผิวของเซลล์ของอวัยวะนี้ โดยในส่วนที่เป็นฐานประกอบไปด้วยมอเตอร์แบบหมุนขนาดเล็กมากตัวหนึ่ง ยึดติดกับสายฟิลาเมนท์ที่เป็นเกลียวบางยาวประมาณ 15 ไมครอน มอเตอร์ของออร์แกเนลล์สามารถที่จะทำงานโดยการหมุนได้โดยมีรอบสูงสุดประมาณ 20,000 รอบต่อนาที หรือ 20,000 rpm (revolution per minute) แต่ใช้พลังงานในการหมุนเพียง 10 -16 วัตต์ และมีประสิทธิภาพในการอนุรักษ์พลังงานได้เกือบร้อยเปอร์เซ็นต์เลยทีเดียว
|
||||||
|
||||||
แฟลเจลลัมถูกสร้างขึ้นมาด้วยความสามารถในการประกอบตัวเองได้ (self-assembly) ของโปรตีนรวมทั้งสิ้น 25 ชนิด วงแหวนตัวหมุนถูกสร้างขึ้นมาจากโปรตีน FliF เป็นส่วนประกอบลำดับแรกที่ถูกประกอบขึ้นมาให้อยู่ติดกับเยื่อหุ้มเซลล์ จากนั้นโมเลกุลของโปรตีนอื่นก็จะประกอบกันขึ้นเป็นวงแหวนวงอื่นๆ ที่อยู่บนโครงสร้างส่วนฐานของมอเตอร์ หลังจากที่มอเตอร์ก่อรูปขึ้นและสร้างจนเสร็จสมบูรณ์แล้ว สายยาวลักษณะคล้ายแส้ที่อยู่ในแฟลเจลลัมก็จะถูกสร้างขึ้นเป็นลำดับต่อมา (หรือเรียกกันว่าแฟลเจลลา (flagellar)) โดยสายแฟลเจลลาจะถูกสร้างขึ้นโดยการเชื่อมต่อกันเป็นพอลิเมอร์ของหน่วยย่อยแฟลเจลลิน (flagellin) จำนวน 20,000 – 30,000 หน่วยเข้าเป็นโครงสร้างที่มีลักษณะเป็นเหมือนท่อเกลียว ซึ่งโมเลกุลแฟลเจลลินจะถูกส่งผ่านมาภายในช่องแคบที่อยู่ในตำแหน่งใจกลางของแฟลเจลลัม โดยส่งออกมาในทิศทางจากภายในตัวเซลล์ไปสู่ทางด้านส่วนปลายของสายแฟลเจลลา |
||||||
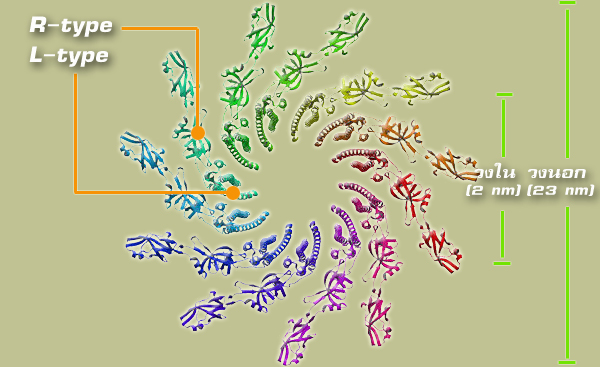
แฟลเจลลาประกอบไปด้วยสายเกลียวจำนวนทั้งหมด 11 สาย โดยในแต่ละสายนั้นจะมีลักษณะการจัดเรียงตัวที่แตกต่างกันออกเป็น 2 แบบ ได้แก่ L-type และ R-type ซึ่งสายเกลียวตั้งต้นจะเป็นแบบ L-type มีความยาวประมาณ 5.27 นาโนเมตร แล้วต่อจากนั้นยาวออกมาจะเป็นสายลักษณะแบบ R-type ซึ่งมีความยาวประมาณ 5.09 นาโนเมตร และสายเกลียวทั้งสองแบบนี้ก็จะจับตัวเข้าด้วยกันเป็นสายเดียวต่อเนื่องกัน ทำให้โครงสร้างของสายแฟลเจลลานั้นมีลักษณะเป็นท่อเกลียวได้ |
||||||
|
||||||
การที่สายแฟลเจลลาสามารถที่จะประกอบตัวเองเข้าเป็นเกลียวได้ก็เนื่องจากการช่วยเหลือของแคพ (cap) ที่อยู่ส่วนยอดสุดของสายแฟลเจลลา โดยแคพมีรูปร่างลักษณะเป็นแบบห้าเหลี่ยม ประกอบไปด้วยแผ่นเพลทห้าเหลี่ยม 1 แผ่นและขาย่อย 5 ขาอยู่ร่วมกันเป็นวง แคพถูกสร้างขึ้นมาจากโปรตีน Flid หรือ HAP2 (hook-associated protein2) โดยที่แคพนี้เป็นกลไกสำคัญสำหรับกระบวนการของการประกอบตัวเองได้ของโมเลกุลแฟลเจลลิน โดยทำหน้าที่จัดเตรียมพื้นที่ยึดติดสำหรับโมเลกุลแฟลเจลลินในแต่ละช่วงเวลาของการสร้างเป็นสายแฟลเจลลา และยังเป็นตัวนำทางระหว่างการยึดติดกันของโมเลกุลแฟลเจลลิน |
||||||
|
||||||
ด้วยคุณสมบัติของการที่โมเลกุลรู้จักว่าจะต้องวางตัวอย่างไรเมื่ออยู่ในระหว่างตัวอื่น ในรูปแบบเชิงโครงสร้างของการก่อร่างสายแฟลเจลลา และการสร้างมอเตอร์แบบหมุนนี้ ทำให้กระบวนการสร้างแฟลเจลลัมโดยความสามารถในการประกอบกันได้นั้นมีความถูกต้องแม่นยำ และเกิดความผิดพลาดน้อยมาก หรืออาจจะปราศจากข้อผิดพลาดเลยก็ว่าได้ |
||||||
|
||||||
แต่สำหรับเซลล์ยูคาริโอต แฟลเจลลัมมีรูปร่างเป็นแท่งทรงกระบอก มีเส้นผ่าศูนย์กลางประมาณ 0.2 ไมครอน โครงสร้างของซิเลียและแฟลกเจลลัมในเซลล์ยูคาริโอตไม่แตกต่างกัน คือประกอบด้วยชุดของไมโครทิวบิล 9 ชุดจัดเรียงตัวเป็นวงกลมอยู่รอบนอก โดยแต่ละชุดประกอบด้วยไมโครทิวบิล 2 แท่ง ส่วนไมโครทิวบิลที่อยู่เป็นเส้นเดี่ยวจะอยู่ตรงกลางอีก 2 แท่ง (เรียกว่ามีลักษณะแบบ 9+2) แต่ละแท่งคู่ของไมโครทิวบิลที่อยู่รอบนอกจะมีแขนของไดนีอีน (dynein arms) ยื่นออกไปสัมผัสกับแท่งคู่ที่อยู่ใกล้เคียงกัน และมีแกนยึด (radial spoke) ยื่นเข้าไปติดต่อกับแท่งคู่ที่อยู่ตรงกลาง โดยในซิเลียจะมีขนาดความยาวประมาณ 10 ไมครอนและมีจำนวนมาก ส่วนแฟลเจลลัมจะมีขนาดความยาวที่ยาวกว่า คืออาจยาวได้ถึง 150 ไมครอนและมีจำนวนน้อย โดยอาจมีเพียงเซลล์ละ 2 - 3 เส้นเท่านั้น |
||||||
| การเคลื่อนไหวของแฟลเจลลัมเพื่อให้เกิดการเคลื่อนที่ของเซลล์โปรคาริโอตนั้น จะมีลักษณะเป็นแบบแกว่งคล้ายไม้พาย แต่สำหรับแฟลเจลลัมที่พบในเซลล์ยูคาริโอตนั้นจะพบว่ามีการเคลื่อนไหวแบบเป็นคลื่นเสมอ ซึ่งเกิดจากการทำงานของมอเตอร์โปรตีนไดนีอีน (dynein) ขนาดประมาณ 50 นาโนเมตร ที่เกาะกับแท่งไมโครทิวบิลคู่หนึ่งไปเกาะกับอีกแท่งคู่หนึ่ง โดยเป็นการเกาะและปล่อยสลับกันไปจนทำให้แท่งไมโครทิวบิล เกิดการเลื่อนที่ในแนวข้างหรือสไลด์ไปมาตามการทำงานของมอเตอร์ไดนีอีน พบว่าในการสไลด์ได้จะต้องใช้ไดนีอีนอย่างน้อยจำนวน 12 โมเลกุล โดยแต่ละครั้งของการสไลด์ทำให้เกิดระยะทาง 8 นาโนเมตร และระยะทางโดยรวมของการสไลด์มากที่สุดเท่ากับ 100 นาโนเมตร ซึ่งในการทำงานได้ของไดนีอีนนี้ต้องใช้พลังงานจากโมเลกุลเอทีพีเพื่อสร้างการเคลื่อนไหวนี้ จึงมีผลทำให้เกิดเป็นการเคลื่อนไหวของแฟลเจลลัมหรือซีเลียในลักษณะเบนโค้งโก่งตัวกลับไปมาได้ (ลักษณะคล้ายคลื่น) จนทำให้เซลล์ยูคาริโอตสามารถเคลื่อนที่ได้
|
||||||









_35.jpg "singleclickstop=[on] header=[นาโนเทคโนโลยีชีวภาพในระบบธรรมชาติ]
body=[กรงนาโนของแชพเพอโรนิน - มอเตอร์ที่ใช้ขับเคลื่อนแฟลเจลลัมของแบคทีเรีย]")
 - มอเตอร์เชิงเส้นของไมโครทิวบิล(ไคเนซิน-ไมโครทิวบิล)]")
]")
 - ช่องเข้าออกเยื่อหุ้มเซลล์ควบคุมด้วยความต่างศักย์ไฟฟ้า(Voltage-gated ion channel)]")



