![]()
![]()
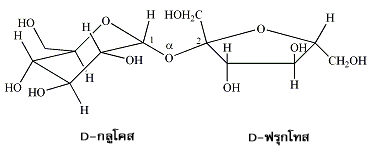
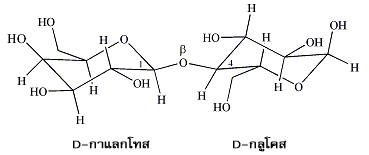
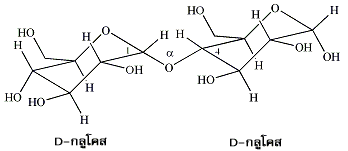
หมู่ไฮดรอกซิลของแอโนเมอริกคาร์บอนในน้ำตาลโมเลกุลเดี่ยวรวมทั้งอนุพันธ์ของมันสามารถเกิดการรวมตัวกับหมู่ไฮดรอกซิลของน้ำตาลอีกตัวหนึ่งที่ตำแหน่ง 1, 2, 3, 4 หรือ 6 ก็ได้ เกิดเป็น แอลฟาไกลโคไซด์ และ บีตาไกลโคไซด์ (α- and ß-glycosides) ส่วนพันธะที่เชื่อมต่อระหว่างแอโนเมอริกคาร์บอนกับแอซีตาลออกซิเจน (acetal oxygen) เป็นพันธะโคเวเลนต์ที่มีชื่อเรียกเฉพาะว่า พันธะไกลโคซิดิก (glycosidic bond) น้ำตาลโมเลกุลคู่ (disaccharides) ประกอบด้วย น้ำตาลโมเลกุลเดี่ยว 2 หน่วย ที่เชื่อมต่อกันด้วยพันธะไกลโคซิดิกซึ่งอาจเป็นชนิดแอลฟาหรือบีตา ตัวอย่างเช่น ซูโครส (sucrose) หรือน้ำตาลทรายซึ่งได้จากต้นอ้อยเป็นน้ำตาลโมเลกุลคู่ที่มีน้ำตาลกลูโคสหนึ่งหน่วยเชื่อมต่อกับน้ำตาลฟรุกโทสหนึ่งหน่วย โดยที่หมู่ไฮดรอกซิลของแอโนเมอริกคาร์บอนในกลูโคสเกิดการรวมตัวกับหมู่ไฮดรอกซิลตรงคาร์บอนตำแหน่งที่ 2 ของฟรุกโทส และพันธะไกลโคซิดิกที่เกิดขึ้นเป็นชนิดแอลฟา
พันธะไกลโคซิดิกแบบแอลฟาและบีตาที่เกิดขึ้นจากการที่น้ำตาลโมเลกุลเดี่ยวตั้งแต่สองหน่วยมาประกอบกัน จะมีความเสถียรและไม่มีการเปลี่ยนโครงรูปกลับไปกลับมาระหว่างรูปแอลฟากับบีตาของน้ำตาลโมเลกุลเดี่ยวตัวนั้นๆ ดังนั้นพันธะไกลโคซิดิกที่เชื่อมระหว่างน้ำตาลแต่ละตัวของน้ำตาลโมเลกุลคู่ (disaccharides ซึ่งมีน้ำตาลโมเลกุลเดี่ยว 2 หน่วย ) คาร์โบไฮเดรตสายสั้น (oligosaccharides ซึ่งมีน้ำตาลโมเลกุลเดี่ยว 3-15 หน่วย ) และคาร์โบไฮเดรตตัวโต (polysaccharides ซึ่งมีน้ำตาลโมเลกุลเดี่ยวมากกว่า 100 หน่วย ) จึงเป็นชนิดแอลฟาหรือบีตาเพียงอย่างใดอย่างหนึ่งเท่านั้น
คาร์โบไฮเดรตส่วนใหญ่ที่พบในธรรมชาติจะอยู่ในรูปของคาร์โบไฮเดรตตัวโตที่มีน้ำตาลโมเลกุลเดี่ยวมาเชื่อมต่อกันอยู่เป็นจำนวนนับร้อย นับพัน ถึงหมื่นแสน น้ำตาลที่มาประกอบเข้าด้วยกันอาจเป็นน้ำตาลชนิดเดียวกัน (homopolysaccharides) หรือต่างชนิดกัน (hetropolysaccharides) ที่ต่างชนิดกันก็ต่างกันเพียง 2-3 ชนิดเท่านั้น และการเชื่อมต่อกันของน้ำตาลโมเลกุลเดี่ยวอาจเป็นการต่อกันแบบ สายยาวที่ไม่มีกิ่งก้านสาขา (linear) หรือสายยาวที่มีกิ่งก้านสาขา (branched) คาร์โบไฮเดรตโมเลกุลใหญ่ที่ประกอบด้วยน้ำตาลโมเลกุลเดี่ยวชนิดเดียวกันทั้งโมเลกุล มักพบว่าทำหน้าที่เป็นเสบียงสะสมอาหารของเซลล์ เช่น แป้งที่สะสมอยู่ในเนื่อเยื่อของพืชและไกลโคเจนที่สะสมอยู่ในเนื้อเยื่อของสัตว์ บางชนิดทำหน้าที่ทางด้านโครงสร้างที่ให้ความแข็งแรง เช่น เซลลูโลสซึ่งเป็นองค์ประกอบที่สำคัญของผนังเซลล์พืช และไคตินซึ่งเป็นองค์ประกอบของเปลือกหุ้มตัวแมลงและปู กุ้ง คาร์โบไฮเดรตโมเลกุลใหญ่ที่ประกอบด้วยน้ำตาลโมเลกุลเดี่ยวต่างชนิดกัน มักพบว่าทำหน้าที่เป็นกันชนในสิ่งมีชีวิต เช่น ในกระดูกอ่อน ผิวหนัง และหลอดเลือด เนื่องจากมีความสามารถในการอุ้มน้ำสูง ส่วนคาร์โบไฮเดรตสายสั้นๆ มักพบที่ผิวเซลล์ในรูปของไกลโคโปรตีนหรือไกลโคลิพิด ทำหน้าที่ในการจับกับโมเลกุลจากภายนอกเซลล์ ที่ทำหน้าที่เป็นสัญญาณ แล้วส่งต่อสัญญาณนั้นเข้าไปภายในเซลล์
แป้งและไกลโคเจนเป็นเสบียงสำคัญของเซลล์ แป้งในพืชมี 2 ชนิด คือ อะไมโลส (amylase) และ อะไมโลเพกติน (amylopectin) ทั้งสองชนิดถูกเก็บสะสมไว้ในส่วนของเหลวในเซลล์ (cytoplasm) สำหรับไว้เป็นเสบียง แป้งมีมากในพืชที่มีหัว เช่น เผือก มัน และในส่วนที่เป็นเมล็ด เช่น ข้าว
ส่วนโครงสร้างของอะไมโลเพกตินประกอบด้วยกลูโคสนับล้านหน่วย มีลักษณะเป็นสายยาวที่มีจุดแตกแขนงแยกออกไปโดยเฉลี่ยทุกๆ 20-30 หน่วยของกลูโคส พันธะไกลโคซิดิกในโครงสร้างของอะไมโลเพกตินเป็นชนิดแอลฟาเช่นเดียวกับอะไมโลส โดยที่พันธะไกลโคซิดิกของโครงสร้างหลักเชื่อมระหว่างแอโนเมอริกคาร์บอนของกลูโคสตัวหนึ่ง กับหมู่ไฮดรอกซิลของคาร์บอนตำแหน่งที่ 4 ของกลูโคสตัวถัดไป แต่ตรงตำแหน่งที่เป็นจุดแตกแขนง พันธะไกลโคซิดิกจะเชื่อมระหว่างแอโนเมอริกคาร์บอนของกลูโคสตัวหนึ่ง กับหมู่ไฮดรอกซิลของคาร์บอนตำแหน่งที่ 6 ของกลูโคสอีกตัว โมเลกุลของไกลโคเจนประกอบด้วยน้ำตาลกลูโคสชนิดเดียวเช่นกัน โครงสร้างของไกลโคเจนมีลักษณะคล้ายกับอะไมโลเพกตินคือมีจุดที่แตกแขนงเป็นกิ่งก้านสาขา แต่จุดแตกแขนงของไกลโคเจนจะถี่กว่า คือพบโดยเฉลี่ยทุกๆ 8 ถึง 14 หน่วยของกลูโคส
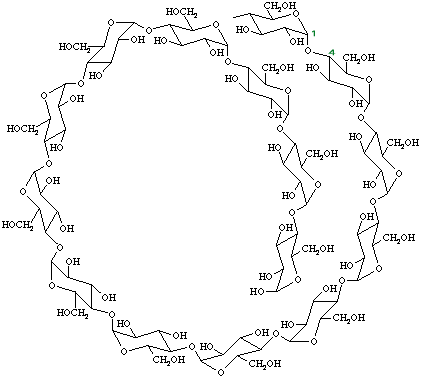
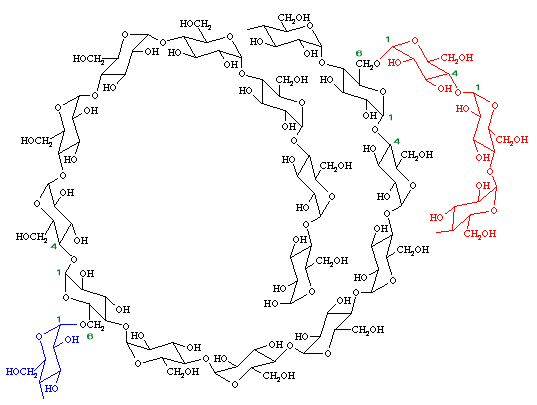
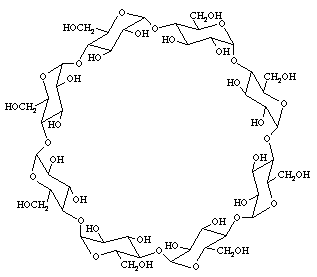
ในธรรมชาติยังมีคาร์โบไฮเดรตที่มีโครงสร้าง 3 มิติเป็นรูปวง ที่ชื่อไซโคลเดกซ์ตริน (cyclodextrin) ซึ่งมีโครงสร้างหลักของสายคาร์โบไฮเดรตเป็นแป้งที่ผ่านการย่อยจนเหลือกลูโคสที่เป็นองค์ประกอบ 6-12 หน่วย โครงสร้างรูปวงเกิดขึ้นโดยอาศัยการทำงานของเอนไซม์มอลโทเด็กซ์ตรินกลูคาโนทรานส์เฟอเรส (maltodextrin glucanotransferase) ของแบคทีเรียที่ชื่อ บาซิลัส มาซีแรนส์ (Bacillus macerans) ในการตัดโมเลกุลของแป้งและเชื่อมต่อคาร์โบไฮเดรตสายสั้นที่ได้จากการย่อยแป้งดังกล่าวเป็นให้เป็นรูปวง โครงสร้างรูปวงนี้มีประโยชน์ในการบรรจุสารโมเลกุลเล็กๆ ที่แบคทีเรียได้รับจากเซลล์เจ้าบ้าน เอาไว้ตรงกลาง และในทางอุตสาหกรรมอาหารเรายังสามารถนำไปประยุกต์ใช้ในการคงสภาพของวิตะมิน สารให้กลิ่น และเคลือบสารที่มีรสขมต่างๆ
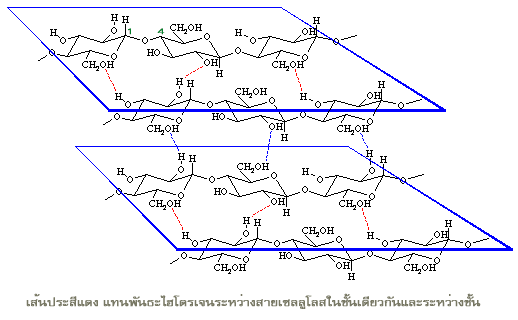
โครงสร้างของเซลลูโลสประกอบด้วยกลูโคสเพียงชนิดเดียวเช่นเดียวกับแป้ง และเป็นชนิดสายตรงไม่มีกิ่งก้านสาขา แต่พันธะไกลโคซิดิกในโมเลกุลของเซลลูโลสจะเป็นชนิดบีตา จึงทำให้กลูโคสที่ต่อกันอยู่ในสายของเซลลูโลสยืดตัวออกในแนวเส้นตรงได้ แต่ละสายของเซลลูโลสมีกลูโคสเชื่อมต่อกันอยู่ ประมาณ 2,000 ถึง 10,000 หน่วย
เส้นใยแต่ละเส้นของพืชซึ่งมีความเหนียวและให้ความแข็งแรงแก่โครงสร้างของผนังเซลล์พืชนั้นประกอบด้วยสายของเซลลูโลส 30 ถึง 40 สาย เรียงตัวขนานกันและอัดตัวกันแน่นเป็นชั้นๆ รวมอยู่กับโมเลกุลขนาดใหญ่อื่นๆ เช่น ฮีมิเซลลูโลส (hemicellulose) เพกติน (pectin) และ ลิกนิน (lignin) โมเลกุลเหล่านี้ทำหน้าที่คล้ายซีเมนต์โบกฉาบเพิ่มความแข็งแกร่งให้แก่ผนังเซลล์พืช
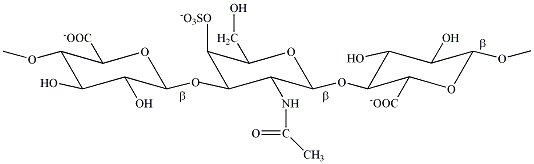
ฮีมิเซลลูโลสและเพกตินเป็นคาร์โบไฮเดรตสายยาว หน่วยย่อยส่วนใหญ่ของฮีมิเซลลูโลสเป็นน้ำตาลไซโลส (xylose) แต่อย่างไรก็ตามในโครงสร้างไม่ได้มีส่วนใดเป็นเซลลูโลสตามชื่อของมันเลย สำหรับเพกตินนั้นมีองค์ประกอบส่วนใหญ่เป็นกรดกาแลกตูโรนิกหรือเกลือของมัน ส่วนลิกนินนั้นเป็นสารอินทรีย์โมเลกุลใหญ่ ที่มีหน่วยย่อยเป็นวงแหวนอะโรมาติกคล้ายเบนซีนที่เชื่อมต่อกับหมู่ต่างๆ ของ มีสัตว์เพียงไม่กี่ชนิดที่มีเอนไซม์ซึ่งสามารถย่อยพันธะไกลโคซิดิกของเซลลูโลสได้ เช่น วัว ควาย กวาง เป็นต้น อันที่จริงที่สัตว์พวกนี้สามารถใช้เซลลูโลสเป็นแหล่งพลังงานได้ก็เนื่องจากว่ามันมีจุลินทรีย์บางชนิดในทางเดินอาหารของมัน จุลินทรีย์เหล่านั้นมีเอนไซม์ที่สามารถย่อยเซลลูโลสให้อยู่ในสภาพที่จะนำเอาไปใช้ประโยชน์ได้ สัตว์จำพวกกุ้ง ปู และแมลงต่างๆ มีกระดองหรือเปลือกนอกเป็นโครงสร้างที่ให้ความแข็งแรง เทียบได้กับส่วนผนังเซลล์ของพืช เปลือกห่อหุ้มสัตว์เหล่านี้มีองค์ประกอบหลักเป็นไคตินซึ่งอยู่รวมกับโปรตีนและแคลเซียมคาร์บอเนต โครงสร้างของไคตินมีลักษณะเช่นเดียวกับเซลลูโลส ประกอบด้วยอนุพันธ์ของน้ำตาล ชนิดที่เป็นน้ำตาลอะมิโนตัวที่ชื่อ N- แอซีทิล-D- กลูโคซามีน (N-acetyl-D-glucosamine) ซึ่งเชื่อมต่อกันด้วยพันธะไกลโคซิดิกแบบบีตา
ไกลโคซามิโนไกลแคนทำหน้าที่เป็นกันชน เนื้อเยื่อเกี่ยวพัน (connective tissues) ในร่างกายของเรา เช่น กระดูกอ่อน เอ็น ผิวหนัง และหลอดเลือด มีคอลลาเจน (collagen) และอีลาสติน (elastin) แทรกตัวอยู่ในช่องว่างระหว่างเซลล์ โดยที่เส้นใยโปรตีนทั้งสองชนิดนี้ฝังตัวอยู่ในของเหลวข้นหนืดคล้ายวุ้น (gel) ที่มีส่วนประกอบหลักเป็น ไกลโคซามิโนไกลแคน (glycosaminoglycans)
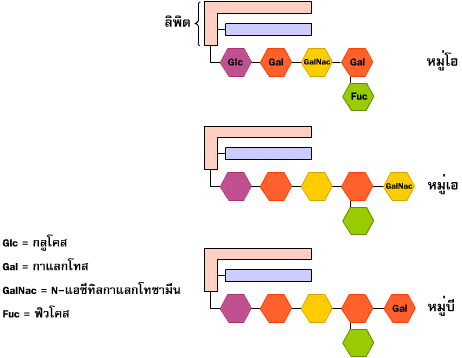
ไกลโคโปรตีนเป็นสารประกอบของคาร์โบไฮเดรตและโปรตีน ในสัตว์จะพบอยู่ที่ผิวเซลล์ และในของเหลวต่างๆ ในร่างกาย คาร์โบไฮเดรตที่พบในไกลโคโปรตีนจะมีน้ำตาลโมเลกุลเดี่ยวเป็นองค์ประกอบประมาณ 2-15 โมเลกุล น้ำตาลที่พบมากในโมเลกุลของไกลโคโปรตีนได้แก่ กาแลกโทส แมนโนส ฟิวโคส N- แอซีทิลกาแลกโทซามีน และ กรด N- แอซีทิลนิวรามินิก หรือ กรดไซอะลิก (sialic acid) โครงสร้างของคาร์โบไฮเดรตมีโอกาสเกิดขึ้นได้หลายแบบต่างๆ กัน ทั้งที่เป็นสายยาวและแบบที่มีกิ่งก้านสาขาโดยที่ความหลากหลายนี้ไม่ถูกกำหนดโดยตรงจากยีนหรือพันธุกรรม แต่เกิดขึ้นจากปฏิกิริยาของเอนไซม์ที่ทำหน้าที่ในการสังเคราะห์คาร์โบไฮเดรตเหล่านั้น เซลล์แต่ละชนิดในร่างกายของเราจึงใช้ประโยชน์จากความหลากหลายของคาร์โบไฮเดรตที่เป็นองค์ประกอบของไกลโคโปรตีนที่อยู่บนผิวเซลล์ในการเป็นเครื่องหมายหรือรหัส เพื่อให้เซลล์แต่ละชนิดจดจำกันได้ หรือเป็นที่ให้โมเลกุลที่จำเพาะมาเกาะเพื่อที่จะเข้าไปในเซลล์ หรือส่งสัญญาณต่างๆ ให้เซลล์รับทราบ คาร์โบไฮเดรตสายสั้นๆ ของไกลโคลิพิดที่เยื่อเซลล์เม็ดเลือดแดงของเรายังมีส่วนทำให้เม็ดเลือดแดงมีความจำเพาะต่อบุคคล ยกตัวอย่างเช่น หมู่เลือดเอ บี โอ คาร์โบไฮเดรตที่เป็นตัวกำหนดหมู่เลือดเหล่านี้จะต่ออยู่กับลิพิดที่อยู่ทางด้านนอกของผนังเม็ดเลือดแดง ผลจากการได้รับเลือดผิดหมู่ ทำให้ระบบภูมิคุ้มกันจำแนกเม็ดเลือดที่ได้รับเข้ามาใหม่นี้ว่าเป็นสิ่งแปลกปลอมนอกร่างกาย และจะทำการตกตะกอนโปรตีนของเม็ดเลือดแดงใหม่นั้น ทำให้เกิดอันตรายถึงแก่ชีวิตได้
|
|||||||||||||||||||||||||||||||||||